


 Email this Article
Email this Article Print
Print Download (PDF)
Download (PDF)
Abstract 
In
the year 2000 the genus Teagueia Luer (Orchidaceae, subtribe
Pleurothallidinae) contained only six species, all epiphytic.
Recently we have discovered 26 unusual new terrestrial species of
Teagueia on four neighboring mountains in the Upper Pastaza
Watershed. All 26 species share distinctive floral and vegetative
characters not found in the six previously described members of
Teagueia, suggesting that all 26 evolved locally from a recent
common ancestor. Each of the Teagueia Mountains hosts 7-15
sympatric species of new Teagueia, suggesting that the
speciation events, which produced this radiation, may have occurred
in sympatric populations.
There
is little overlap in the Teagueia species hosted by the
mountains, though they are separated by only 10-18 km. This is
difficult to explain in light of the high dispersal ability of most
orchids. Surprisingly, the species appear not to be habitat
specialists. In forest above 3100 m, population densities of these
new Teagueia are higher than those of any other genera of
terrestrial flowering plants. In the paramos of these mountains they
are the most common orchids; they reach higher elevations (3650m)
than any other pleurothallid orchid except Draconanthes aberrans.
While most species occur
at high densities, three species are known only from single examples.
These species must have population centers on nearby unexplored
mountains, suggesting that we are only tapping the surface of this
radiation. The existence of such a rich and completely unexpected
flora, with such high beta diversity, poses serious challenges for
conservation.
Keywords:
Beta diversity, sympatric speciation, dispersal
Resumen
En
el año 2000 el genero Teagueia Luer (Orchidaceae,
subtribu Pleurothallidinae) tenia únicamente seis especies,
todas epifitas. Desde esta fecha hemos descubierto 26 nuevas especies
de este género, todos terrestres, en cuatro cerros conjuntos
de la cuenca alta del Rio Pastaza. Todas las 26 especies comparten
unas características distintas que no tienen las seis especies
anteriormente descrito, sugierendo que todas las 26 han evolucionado
localmente desde un ancestro común en el pasado reciente. Cada
una de los cerros de los Teagueia tiene 7-15 especies
simpatricas de estas nuevas especies de Teagueia sugieren que
los procesos de especiación en esta radiación pueden
haber ocurrido mientras las especies eran simpatricas.
Hay
pocas especies compartiendo entre estos cerros, a pesar de que su
separación es no mas que 10-18 km. Es difícil explicar
esta situación, porque las orquídeas poseen altas
capacidades para dispersión de semillas sobre largas
distancias. Parece que las especies no son especialistas en hábitats
específicos. En bosques sobre los 3100 m, las densidades de
estas nuevas especies de Teagueia son mas alta que en
cualquier otro genero de plantas avanzadas. En los paramos de estos
cerros las especies de Teagueia son las orquídeas mas
comunes; llegan a alturas sobre (3650 m), que cualquier otra especies
de orquídea de subtribu Pleurothallidinae excepto Draconanthes
aberrans.
Mientras
la mayoría de las nuevas especies de Teagueia existen
en densidades altas, tres de ellas están conocido únicamente
por un solo ejemplar. Estas tres especies deben tener sus centros de
población en otros cerros de la cuenca, indicando que nuestro
conocimiento sobre radiación, aún es escaso. La
existencia de una flora tan rica e inesperada, con una diversidad
beta tan alta pone serios desafíos para la conservación.
Introduction
There have been many localized species-level plant radiations in South America, the most famous of which is probably the Scalesia radiation discovered by Charles Darwin in the Galapagos Islands of Ecuador. That radiation consists of fifteen species of Scalesia, spread out over about thirty thousand square kilometers, with a maximum of six sympatric species per island (Valencia et al 2000). I here report the discovery of a completely unsuspected species radiation, far more dramatic than the Scalesia radiation in terms of number and density of species, in the high mountains of the Upper Pastaza Watershed in the eastern Andes of Ecuador. This radiation consists of about 26 new closely related species of the genus Teagueia (Orchidaceae), growing on four adjacent mountains just 10-12 km apart, with up to 15 species sympatric on a single mountain.
The Upper Pastaza Watershed (here defined as the watershed of the Rio Pastaza from 900 m upward) is the deepest and straightest valley in the eastern Andes of Ecuador, and it serves as a major conduit for airflow between Amazonia and the cold dry Interandean Valley. The interaction of this airflow with the irregular topography creates many unusual microclimates, and this has led to the evolution of many endemic species of plants. The watershed is seldom appreciated as a center of endemism, but in fact the number of vascular plant species strictly endemic to the Upper Pastaza Watershed exceeds the number endemic to the Galapagos Islands. About half of these endemic species are orchids. Though the area has a long history of botanical exploration, beginning with Richard Spruce in 1857, the higher mountains of the watershed (above 3000 m) are nearly inaccessible and are virtually unknown botanically.
Methods
This work is part of a larger study to map the distributions of Teagueia Luer, Lepanthes Sw., and certain other pleurothallid orchid genera in the Upper Pastaza Watershed, concentrating on a long strip 8 km wide centered on the Rio Pastaza, beginning in Puyo and ending above Baños. Sampling has been ongoing for six years. Sampling is limited by the steepness of the terrain, but all major ridgelines and most minor ones have now been sampled to about 2400 m. Six of the seven highest ridge systems have now been sampled to 3100m or beyond. Locations since 2001 have been georeferenced using a GPS, and voucher collections for each species of important genera (Lepanthes, Teagueia, Masdevallia, Dracula, Trichosalpinx, Scaphosepalum, Porroglossum) are stored in alcohol. At the end of this study these collections will be distributed to QCNE and QCA. The new orchid species discovered during the course of this project have been described in collaboration with Dr C. A. Luer (Luer 1998, 1999, 2000, 2002 and in press) and Dr. Calaway Dodson (in press).
The discovery of the Teagueia radiation in the Upper Pastaza Watershed was a fortuitous byproduct of this mapping work.
Results
Twenty-six new species of Teagueia Luer were discovered on four adjacent mountains in the Upper Pastaza Watershed, more than quadrupling the size of the genus. All are long-repent, unlike the caespitose or short-repent species previously known, and all share several other features (discussed below) that are unusual or absent in the previously known species.
These new long-repent species show considerable variation both within and between species (Figure 1). They vary significantly in leaf size, ranging from 5-8 mm in the smallest species to 25-35mm in the largest species. The leaf texture varies greatly between species, and also varies somewhat within species; some species have smooth leaves but others have extremely pusticulate upper leaf surfaces. One species has erose leaf margins. Inflorescences are very similarly shaped in most species. The flowers vary greatly in size and shape, though the basic structure is quite similar in all these long-repent species. Colors range from white to yellow to red to black. Many of these long-repent species are polymorphic, having a dark purple form and a yellow form; these colour forms are often found intermixed, as in several other pleurothallid genera (notably Brachionidium, pers. obs).

Figure 1. Examples
of variation in Pleurothallidinae.
Figure 1. Ejemplos de variacion en los Pleurothallidinae.
Figure 1. Ejemplos de variacion en los Pleurothallidinae.
The new long-repent species are found on four adjacent mountains in the Upper Pastaza Watershed: Cerros Mayordomo, Añangu, Negro, and Candelaria (Figure 2). Cerro Mayordomo, north of the Rio Pastaza , hosts seven of the long-repent Teagueia species. Ten kilometers to the west, Cerro Añangu hosts two of the long-repent species found on Cerro Mayordomo and five additional long-repent species. Opposite Cerro Añangu, on the south side of the Rio Pastaza, Cerro Negro hosts 9 long-repent Teagueia species, none of which are found on the north side of the Pastaza. Also on the south side of the Rio Pastaza and directly across from Cerro Mayordomo, Cerro Candelaria hosts fifteen of these new long-repent Teagueia species, including all of the Cerro Negro species, and six additional species unique to Cerro Candelaria. All of these sites are within 18 km of each other. In addition, one population of long-repent Teagueia species has been found 90 km to the south of the Rio Pastaza.
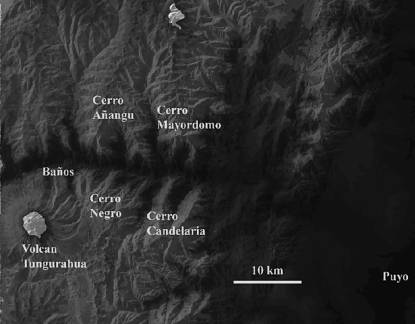
These long-repent Teagueia species are found above 2800-3100 m on each of these four mountains. Their upper elevation limit appears to be 3650 m; above this elevation they were apparently absent on Cerro Candelaria. Most species grow terrestrially in thick moss in deep shade in cloud forests. Most species can be found less commonly growing epiphytically within a meter or two of the ground. The epiphytic habit is more common at higher elevations. Above 3400 m on Cerro Candelaria there are patches of paramo, which often freeze at night, and many of the Teagueia species of that mountain (including several that grow in deep shade at 3100 m) grow in full sun in these paramo patches, often rooting in sphagnum moss.
On all the mountains where long-repent Teagueia species have been found, they are among the most common higher plants, and by far the most common orchids. Their lower altitudinal limit is usually extremely well defined; below a certain point there are no Teagueia plants whatsoever, and above that point they suddenly become abundant. At elevations of 3100 m to 3400 m, densities on the order of five individuals per square meter are common. In a circle of diameter 5m one can normally find three to five species of Teagueia growing together. Some hybridization appears to take place.
The first four long-repent species that were discovered in the course of this work were published in Luer (2000). Descriptions of the more recently discovered species are in preparation.
Discussion
The genus Teagueia Luer (Orchidaceae) was established to contain six unusual species of the subtribe Pleurothallidinae (Luer 1991). Some of these species had been originally placed in the genus Platystele (Luer 1990), where they did not fit very well. Three of these original Teagueia species are Ecuadorian, and three are Colombian. All six species are caespitose or nearly so, and all six have long-tailed sepals and disproportionately small petals. The lip of all six species has an orifice formed by the folded edges of the middle lobe of the lip, and also has a swelling or callus just below the column tip. Most species bear inflorescences with all flowers opening simultaneously.
The 26 new species of Teagueia discovered during the present study all share certain characters not found in the six previously known species. All are long-repent instead of caespitose or short-repent, and none have a swelling or callus on the lip beneath the column. Most have broad sepals without long tails, and all have successively flowered inflorescences rather than the simultaneously flowered inflorescences of most of the previously known species.
The sharing of features that are unusual or absent in the six previously known Teagueia species strongly suggests that all 26 new species evolved from a recent common ancestor. Their extremely limited geographical distribution also suggests a recent origin for this clade. The limited distribution may be an artefact of the inaccessibility of much of the eastern Andes, but it seems likely that they are genuinely absent from the well-studied Papallacta-Baeza-Tena corridor 110 km to the north of the Rio Pastaza, and they are also probably genuinely absent from the well-known Cuenca-Gualaceo-Limon corridor, 180 km to the south of the Rio Pastaza.
One may speculate that the development of the long-repent habit was the key innovation that opened up new niches for this clade. In their extremely wet habitat, competition from fast-growing mosses and liverworts is extreme, and the long-repent habit may allow these species to keep their growing points above the moss while maintaining contact with a humid substrate. It is likely that this innovation was accompanied by a pollinator switch, as indicated by the structural differences between the lips of the long-repent species and those of the "normal" species. The order of these developments is not clear; the pollinator switch may have provided the reproductive isolation needed to permit the fixation of the long-repent habit, or the long-repent habit may have opened up new habitats with a new pollinator regime, leading to the switch.
In any case, the radiation of this clade does not seem to be driven by adaptations to a variety of specific habitats. Many long-repent species typically grow together in the same patch of moss, and most species seem tolerant of a broad range of environmental factors. The radiation of the clade is more likely driven by specialization onto different pollinator niches. One species, T. cymbisepala, has tubular orange flowers and may be hummingbird-pollinated. Small dipterans have been seen visiting some of the other species, though actual pollinization has not been observed. The variety of colours, sizes, and shapes among sympatric species strongly suggests that they each use a different set of pollinators. However, the presence of occasional hybrids suggests that there is some slight overlap in pollinator sets.
It is difficult to explain certain aspects of the distributions of the long-repent Teagueia species. Most unusual is the abrupt transition in the cloud forest between the Teagueia zone and the non-Teagueia zone. Teagueia species change from absent to abundant in the space of a meter or two. This is not a by-product of simple colonial growth, because the boundary line is formed by many different species of Teagueia growing together. All individuals seem to respect the same invisible line in the forest. This suggests that the Teagueia species are all responding to some specific factor in the soil. The obvious candidate for such a factor is a mycorrhizal fungus, since it is known that orchid seed germination depends on the presence of a suitable fungus. There is evidence that species of some other pleurothallid genera, such as Lepanthes, require a genus-specific symbiont fungus (pers. obs.). Perhaps the same is true of Teagueia species, and the invisible line in the forest may be the growth front of a very large mycorrhizal fungus.
Another difficult aspect of Teagueia distribution is the striking difference between the Teagueia floras of each of the four adjacent mountains in the study area. Orchid seeds are the smallest of all flowering plant seeds and are easily dispersed by wind. There can be no doubt that seeds of all Teagueia species cross the valleys that separate these four mountains. Most other orchid species freely cross the Rio Pastaza, and most pleurothallid orchid distributions in the study area form north-south bands related to precipitation (which decreases from east to west). These bands almost always cross the easterly-flowing Rio Pastaza. Yet the Rio Pastaza appears to be an impenetrable barrier for Teagueia species. It is worth noting that the long-repent Teagueia species are the only pleurothallid orchids in the area that are primarily terrestrial. Perhaps some biotic or abiotic soil factor differs from one side of the Rio Pastaza to the other. We suspect that mycorrhizal fungi may be that factor. Some terrestrial mycorrhizal fungi in the temperate rain forests of northwest US are rodent-dispersed. If the mycorrhizal fungus needed by the Teagueia species is rodent dispersed, then the deep canyon of the Rio Pastaza could well form a barrier to its dispersal, and there may be significant differences between the terrestrial mycorrhizal fungi north and south of the Pastaza. It may be that the Teagueia species adapted to the northern fungus cannot germinate on the southern fungus, and vice versa. As a test of this possibility, DNA studies of Teagueia mycorrhizal fungi are planned.
Similar though much smaller local radiations are known in Lepanthes (5 sympatric closely related species on Cerro Abitagua, four of them endemic to Cerro Abitagua; pers. obs.) and Trichosalpinx subgenus Pseudolepanthes (six closely related species on Cerro del Torra in Depto. de Choco, Colombia, four of them endemic to Cerro del Torra; Luer 1997). The Teagueia radiation described here is apparently the most diverse local radiation yet known among South American orchids, and even so, we are probably underestimating its size. Three of these 26 species are known only from single individuals. These three species must have their principal populations elsewhere, hidden in some as-yet-unexplored mountains in the Upper Pastaza Watershed. It is likely that many more species will turn up when these mountains are finally reached.
Acknowledgements
My students Andy Shepard and Pailin Wedel discovered the Cerro Negro species discussed here. This work has been supported by a series of generous grants from John and Ruth Moore to the Population Biology Foundation, and by several grants from the San Diego County Orchid Society, a grant from the Oregon Orchid Society, and grants from the Andean Study Programs, Glenn and Marsha Staats, Alyssa Roberts and Richard Bozek, and Cherise Miller and Kent Udell.
References
Luer, C. A. 1990. Icones Pleurothallidinarum VII. Systematics of Platystele (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 38:1-133.
Luer, C. A. 1991. Icones Pleurothallidinarum VII. Systematics of Lepaanthopsis, Octomeria Subgenus Pleurothallopsis, Restrepiella, Restrepiopsis, Salpistele, and Teagueia (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 64:105-113.
Luer, C. A. 1997. Icones Pleurothallidinarum XV. Systematics of Trichosalpinx (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 64:105-113.
Luer, C. A. 1998. Icones Pleurothallidinarum XVII. Systematics of Subgenus Pleurothallis Section Abortivae, Section Truncatae, Section Pleurothallis Subsection Acroniae, Subsection Pleurothallis, Subgenus Dracontia, Subgenus Unciferia (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 72:103-120.
Luer, C. A. 1999. Icones Pleurothallidinarum XVIII. Systematics of Pleurothallis Subgenus Pleurothallis Section Pleurothallis, Subsection Antenniferae, Subsection Longiracemosae, Subsection Macrophyllae-Racemosae, Subsection Perplexae,, Subgenus Pseudostelis, Subgenus Acuminatia (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 76:139-147.
Luer, C. A. 2000. Icones Pleurothallidinarum XX. Systematics of Jostia, Andinia, Barbosella, Barbodria, and Pleurothallis Subgenus Antilla, Subgenus Effusia, Subgenus Restrepioidia (Orchidaceae). Monographs in Systematic Botany from the Missouri Botanical Garden, 79:1-4, 117-140.
Luer, C. A. 2002. Icones Pleurothallidinarum XXIV. A First Century of New Species of Stelis of Ecuador Part One. Monographs in Systematic Botany from the Missouri Botanical Garden, 88:89-94.
Valencia, R.; N. Pitman; S. Leon-Yanez & P. Jorgensen, eds. 2000. Libro Rojo de las Plantas Endemicas del Ecuador 2000. Herbario QCA, Pontificia Universidad Catolica del Ecuador, Quito.