Introducción

El considerar el número de individuos en la
evaluación del estado de conservación de las especies
es esencial debido a las consecuencias genéticas y
demográficas que presentan las especies con poblaciones de
tamaño pequeño (Watkinson & Harper 1978; Goodman
1987; Carulli & Furbrothers 1988; Menges 1991). Se hipotetiza que
la vulnerabilidad a los eventos ambientales, demográficos y
genéticos al azar son aún más fuertes para las
especies con poblaciones pequeñas. De allí que, se
piensa que las especies raras (en términos de abundancia) son
más propensas a la extinción (MacArthur & Wilson
1967; Terborgh & Winter 1980, Nitecki 1984; Shaffer & Samson
1985; Soulé 1987; Lande 1988; McKinney 1997). En el caso
específico de las plantas, los estudios empíricos
sugieren que las poblaciones pequeñas podrían tener
consecuencias en los procesos ecológicos tales como
polinización (Feinsinger et al. 1991; Kunin 1993; Ågren
1996; Hendrix & Kyhl 2000), y esto podría influenciar en
los procesos evolutivos de la especie (Kunin 1997; Matthies et al.
2004). Cuando las especies de plantas son encontradas en bajo número,
es posible también que presenten rangos geográficos
restringidos (Main 1981; Reveal 1981; New 2000; pero ver Blackburn &
Gaston 1997; Gaston 1997), los cuales son el resultado de
requerimientos ecológicos también restringidos o
específicos. En este sentido, se piensa que los modelos del
nicho ecológico son claves en estructurar la abundancia y la
distribución geográfica de las especies. Por ello,
aunque los factores abióticos no son los únicos que
determinan el nicho ecológico de una especie dada, estos
pueden ser mapeados en áreas geográficas específicas,
y por lo consiguiente son comúnmente usados en los modelos
para determinar la distribución de especies (Carpenter et al.
1993).
En 1998,
Ecuadendron
acosta-solisianum
D. A. Neill (Fabaceae), fue descrito
como un género monotípico endémico de las
Tierras Bajas del oeste del Ecuador (Neill 1998). Esta especie no
solo es el único representante arbóreo endémico
a nivel de género, sino que además esta catalogado como
En Peligro en la naturaleza de acuerdo al Libro Rojo de las plantas
endémicas del Ecuador (Valencia et al. 2000). En aquella
primera descripción Ecuadendronfue
reportado con un número total de individuos adultos menor a
100, en un rango elevacional de 300 - 350-m snm restringido a
bosques húmedos y muy húmedos de la Costa ecuatoriana.
Las poblaciones reportadas llegaban a tres, repartidas entre las
provincias Esmeraldas, Guayas y Cañar (Neill 1998). Sin
embargo, dado que poblaciones adicionales fueron encontradas en las
provincias Guayas y Azuay, el presente estudio tiene como objetivo
utilizar el tamaño poblacional y la distribución
espacial real y potencial como parámetros para actualizar el
estado de conservación de Ecuadendron.
También se estimará la pérdida del hábitat
para esta especie, en vista de que su bajo número de
individuos y su distribución geográfica restringida
podrían deberse a agentes extrínsecos, tales como, las
actividades antropogénicas que han ejercido presión
sobre los recursos naturales por décadas en esta región.
Materiales y Métodos
Especie en estudio
Ecuadendron
acosta-solisianum,
es un árbol de dosel de aproximadamente 30m de alto
cercanamente relacionado con Macrolobium
y Brownea
en la subfamilia Caesalpinioideae (Herendeen et al. 2003). Su corteza
es café claro y su madera es densa y dura en la madurez. Sus
hojas son enteras paripinnadas con pulvínulo basal. La
inflorescencia es péndula, con ~50 flores resupinadas y puede
llegar a medir hasta 2 m de largo, esta resupinación de las
flores sugiere polinización por murciélagos (Neill
1998). Las semillas son grandes (2 - 3.0cm x 2.5 -
5.0cm), sin arilo ni endospermo y se presentan generalmente de 4 -
9 por legumbre. Se desconoce el dispersor aunque el tamaño de
la semilla sugiere dispersión por roedores.

Figura 1: Inflorescencia péndula de Ecuadendron
acosta-solisianum. Sus estructuras florales sugieren polinización por
murciélagos
Area de estudio
Este estudio fue realizado en cinco cerros de la Reserva
Ecológica "Manglares-Churute" (02°24'S,
79°40'W), perteneciente a la provincia Guayas, y en ocho
haciendas en las cercanías del poblado de Manta Real, (02°33'
S, 79°21'W), provincia Azuay y Cañar
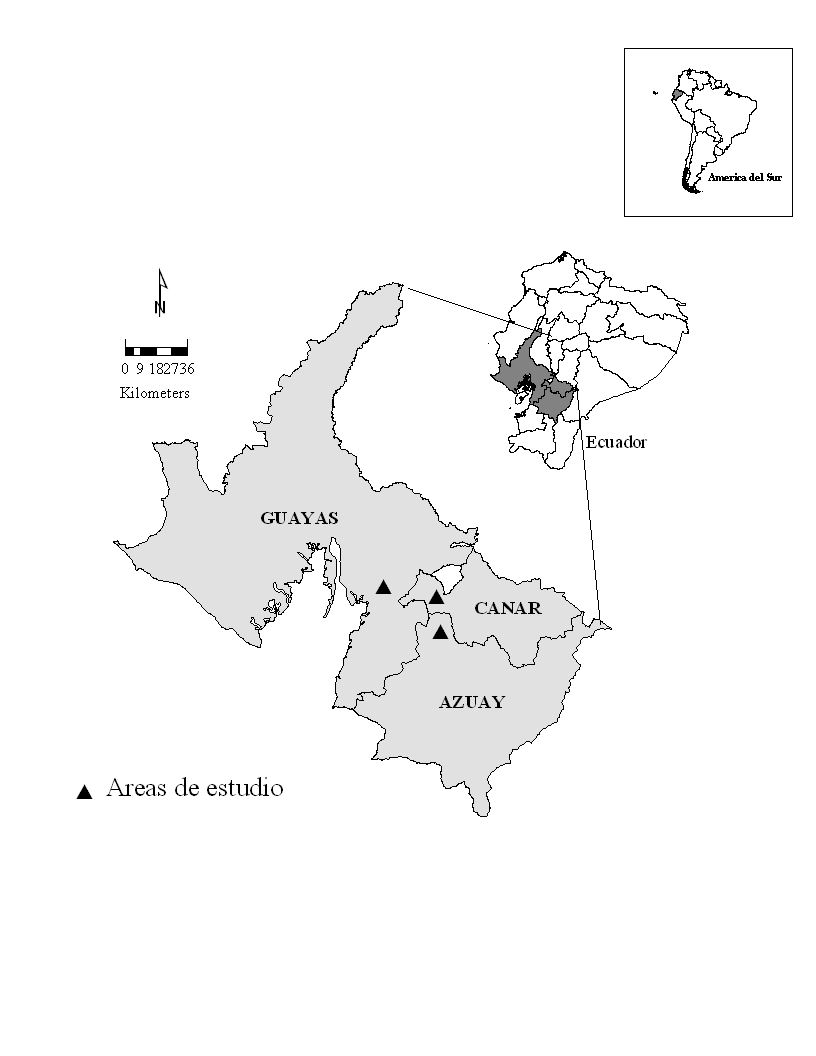
Figura 2: Ubicación de las
áreas de estudio. Cerros de la Reserva Ecológica Manglares-Churute, Provincia Guayas, y propiedades en las provincias Azuay y Cañar alrededor del poblado de Manta Real. Los cerros
de la R. E. Manglares-Churute correspondieron a: cerro Masvale y Pancho Diablo, considerados como
Churute 1, y Perequeté Grande, Simalón y El Mate, considerados como Churute 2
Los cerros de la Reserva Ecológica
Manglares-Churute (REMCh) correspondieron a: cerro Masvale, Pancho
Diablo, Perequeté Grande, Simalón y El Mate. De éstos
el pico más alto correspondió al cerro Pancho Diablo
(727m). Todos ellos son parte de "La Cordillera de Churute"
que recibe la influencia de las subcuencas del Bulubulu-Taura, en el
norte, y Churute, en el sur. La precipitación anual varía
entre 1000 y 1500mm (Mejía et al. 1990), con concentraciones
de lluvia entre Enero y Abril. Se estima una temperatura media anual
de 25.6º C. La humedad mínima, media y máxima se
ha registrado como 70%, 80%, y 85%, respectivamente. Los tipos de
vegetación corresponden a Bosque Semideciduo de Tierras Bajas
de la Costa y Bosque Siempreverde de Tierras Bajas de la Costa
(Sierra 1999). Sobre los 450-m snm, los cerros Masvale, Pancho Diablo
y Simalón se caracterizan por presentar una bruma densa y una
garúa persistente (INEFAN - FUNDACION NATURA /
ECOLAP/USFQ 1999).
Las haciendas en el
sector de Manta Real se encuentran entre un rango altitudinal de 300m
- 1105m, distribuidas entre un complejo de montañas con
uno de los más grandes remanentes de bosque al occidente de
los Andes. La temperatura anual varía entre 18˚ y 22˚
C (Factos 2000). La estación lluviosa va de Diciembre a Abril.
La vegetación de esta área corresponde a Bosque
Siempreverde de Tierras Bajas de la Costa y Bosque Siempreverde
Piemontano de la Costa (Sierra 1999).
Todas estas áreas
pertenecen al complejo "Darién-Chocó-Western
Ecuador" considerado como uno de los mundialmente conocidos
"hotspots" (Mittermeier 1999; Myers et al. 2000).
Diseño del estudio
El trabajo de campo se condujo desde Junio hasta
mediados de Agosto de 2003. Dos de los cinco cerros de la REMCh
(Masvale y Pancho Diablo) fueron tratados como Churute 1, y el resto
de cerros (Perequeté Grande, Simalón y Mate) como
Churute 2. Las propiedades privadas en las cercanías de Manta
Real comprendieron la tercera área de estudio.
Para estimar el tamaño
de la población de Ecuadendron,
establecí transectos variables (Foster et al. 1998;
Peña-Claros 2003) a lo largo de un rango total altitudinal de
20m - 1105m [[Tabla
1]], considerando las tres áreas
de estudio juntas. Este método es una técnica de
muestreo flexible que permite fijar un número de individuos
indistintamente del tamaño del área. Para este estudio,
utilicé 50 individuos adultos por transecto como estándar
para delimitar el largo del transecto. Los transectos empezaron a
20-m snm en Churute 1 y en Churute 2, y a 300-m snm en Manta Real. A
lo largo de cada transecto, primero busqué los individuos
adultos en una distancia de hasta 500-m desde la línea central
del transecto. El muestreo completo en cada transecto se llevó
a cabo por varios días.
Para predecir la
distribución espacial potencial de Ecuadendron
acosta-solisianum
apliqué el modelo del nicho ecológico DOMAIN®
1.4 (Carpenter et al. 1993), el cual mapea áreas de similitud
y produce patrones de distribución usando registros de
ocurrencia y un limitado número de atributos biofísicos
(Carpenter et al. 1993, Loiselle et al. 2003). Para este modelo,
utilicé 142 puntos georeferenciados de presencia de la especie
y cuatro variables bioclimáticas, datum WGS84, a 30 segundos
de resolución espacial, obtenidas de la base de datos
WorldClim 1. 3 (Hijmans et al. 2004). Las variables bioclimáticas
escogidas representan una combinación de temperatura,
precipitación y humedad en diferentes épocas del año,
y correspondieron a: 1) Temperatura promedio del cuarto más
húmedo, 2) Temperatura promedio del cuarto más seco, 3)
Precipitación del cuarto más cálido, y 4)
Precipitación del cuarto más frío. Un cuarto
representa un período de tres meses. El modelo predictivo
final fue preparado utilizando las herramientas del programa de
ESRI®, ArcMap™ 8.2
Análisis de datos
Para estimar el área
muestreada se usó las herramientas del ESRI®
ArcMap™ 9.0 creando un polígono con los
puntos georeferenciados más externos de cada transecto. Para
el modelo de distribución potencial se convirtió el
mapa predictivo DOMAIN® en un mapa binario asignando
nuevos valores a todas las celdas. Así, las celdas con valores
< 95 fueron reclasificadas como 0, equivalente a sitios no aptos
para el desarrollo de la especie, y las celdas con valores de 95 -
100 fueron reclasificadas como 1 (sitios apropiados). La reducción
del hábitat de Ecuadendron
fue estimada con una herramienta para calcular área en
ArcView 3.2 (Zhou 2001).
Resultados y Discusión
Tamaño de la población
Un total de 2,692
individuos de Ecuadendron
fueron muestreados, 163 de los cuales correspondieron a individuos
adultos encontrados en ocho de los 16 transectos muestreados, es
decir, en un transecto en Churute 1, en dos transectos en Churute 2,
y en cinco transectos en Manta Real. En todas las áreas de
estudio, Ecuadendron
estuvo presente en densidades < 1 individuos adultos ha-1.
La densidad de individuos adultos fue mayor en el área de
Manta Real, y Churute 2 tuvo una densidad de adultos más alta
que Churute 1, aunque el área muestreada de Churute 1 fue
mayor que la de Churute 2. Para Manta Real, este resultado contrasta
con la descripción hecha por Foster (1992) quien reporta a
esta especie (identificada como Browneopsis)
como dominante bajo los 400-m snm. Esta observación responde a
una evaluación rápida del área hecha hace más
de doce años. Para Churute 1 y Churute 2, no existen
referencias con las cuales se puedan hacer comparaciones, solo
inventarios parciales que reportan 25 especies de árboles
maderables pertenecientes a las familias Bignoniaceae, Fabaceae,
Lauraceae, Rhizophoraceae y Sapotaceae, que no mencionan datos sobre
frecuencias (INEFAN - FUNDACION NATURA / ECOLAP/USFQ 1999). La
baja densidad de Ecuadendron
podría más bien deberse a las constantes actividades
antropogénicas en estas áreas, aunque la co-existencia
de especies de árboles en bajas densidades parece ser un
patrón común en las comunidades de bosques tropicales
con alta diversidad y alto endemismo (Cody 1986; Gentry 1986; Pitman
et al. 1999).
Distribución potencial
El modelo distribucional
sugiere una presencia discontinua de la especie en 129,164 ha de
bosque al oeste del Ecuador. Los lugares aptos para el
establecimiento de la especie se extienden más allá del
suroeste de los límites de la provincia Esmeraldas y el
noroeste de la provincia Guayas pasando por la provincia de Manabí
(Fig. 3). Su presencia se extiende hacia
la provincia Manabí donde los tipos de
vegetación corresponden a:
Bosque Siempreverde Piemontano de la Costa, Bosque Siempreverde
Piemontano de la Cordillera de la Costa, Bosque Siempreverde Montano
Bajo de la Costa, Bosque Deciduo de Tierras Bajas de la Costa, Bosque
Semideciduo de Tierras Bajas de la Costa, y Bosque Nublado Montano
Bajo (Sierra 1999)]].
Los tipos de vegetación de estas potenciales nuevas áreas
de ocurrencia corresponden a: Bosque Siempreverde Piemontano de la
Costa, Bosque Siempreverde Piemontano de la Cordillera de la Costa,
Bosque Siempreverde Montano Bajo de la Costa, Bosque Deciduo de
Tierras Bajas de la Costa, Bosque Semideciduo de Tierras Bajas de la
Costa, y Bosque Nublado Montano Bajo (Sierra 1999).
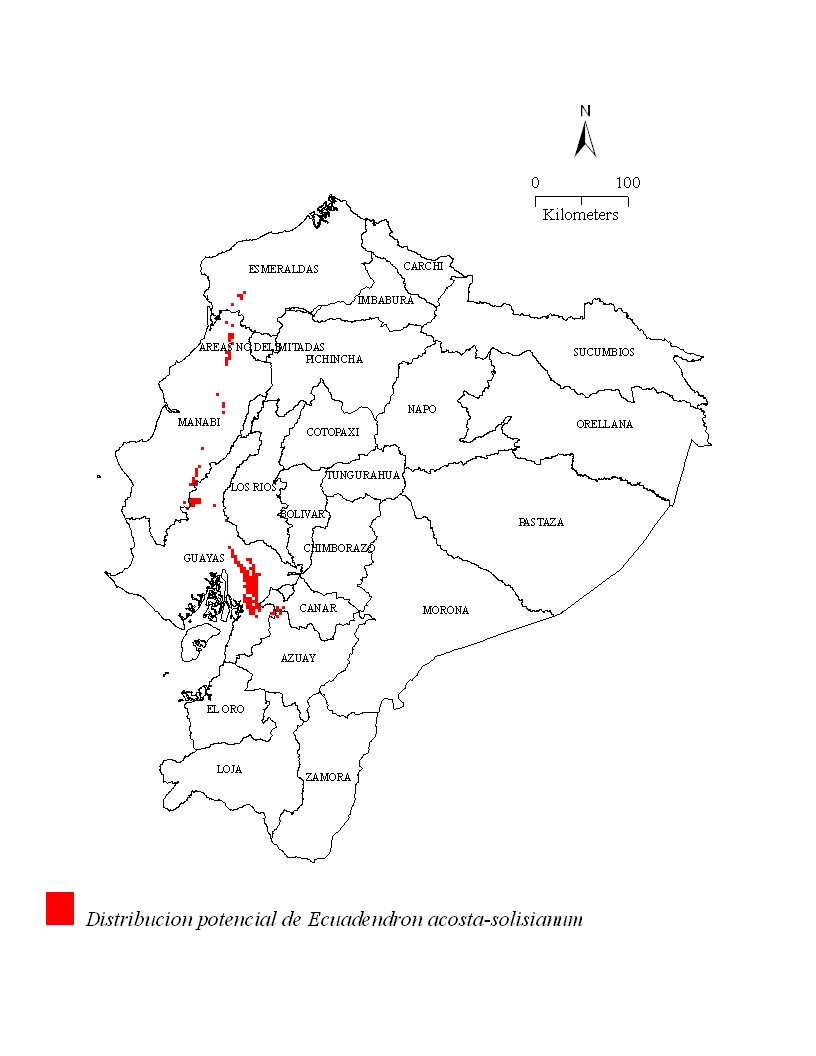
Figura 3: Distribución potencial de
Ecuadendon acosta-solisianum de acuerdo al modelo
predictivo DOMAIN®.
Reducción del hábitat
Se estima que la especie
se encuentra presente en sólo 13,328 ha de las 129,164 ha de
occurrencia representadas en el modelo del nicho ecológico
(Fig. 4). Las áreas remanentes corresponden principalmente a
los bosques al norte de la provincia Esmeraldas. Los causantes más
comunes de la pérdida del hábitat en la provincia Azuay
y Cañar incluyen la conversión de tierra a
plantaciones, la extracción local actual de especies
maderables, así como también el impacto de
explotaciones previas de palmas del género Geonoma,
Jessenia, Pholidostachys,
y miembros de las familias Bombacaceae, Lauraceae and Meliaceae
(Factos 2000). En la provincia Guayas, también se dió
la extracción local antes del establecimiento de la Reserva
Ecológica Manglares-Churute en 1979 (INEFAN-FUNDACION
NATURA/ECOLAP/USFQ 1999).
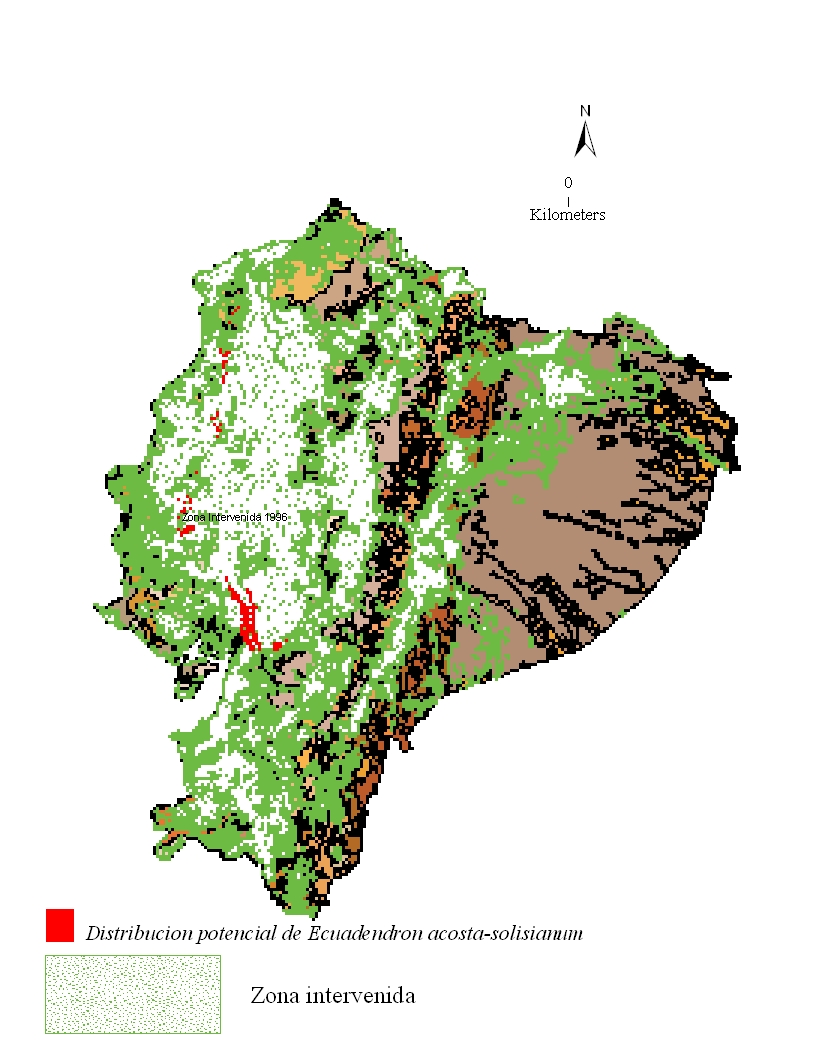
Figura 4: Pérdida del hábitat de Ecuadendron
acosta-solisianum. De norte a sur, se estima que la especie se mantiene en 13,328 ha de
bosque en las provincias Esmeraldas, Manabí, Guayas, Azuay y Cañar, lo cual corresponde a
apenas el 10% de su distribución potencial
Conclusiones
Este estudio constituye
el primer intento en caracterizar las poblaciones de
Ecuadendron
acosta-solisianum
en la naturaleza después de
que fuera taxonómicamente descrita en 1998. Esta
especie fue encontrada rara en la
naturaleza debido a su bajo número de individuos y a su
distribución geográfica restringida, aunque se
registraron nuevas poblaciones y su rango elevacional de presencia se
amplió de 300 - 350-m snm a 170 - 600-m snm de un
rango total muestreado de 20 - 1105-m snm.
La presencia de
Ecuadendron
en menos de un individuo reproductivo por hectárea y su
distribución en pocos remanentes de bosques en el occidente de
Ecuador, puede responder a factores inherentes a su propia biología.
Además, las especies raras parecen ser una característica
común en las regiones con alto endemismo como las Tierras
Bajas del oeste de Ecuador (Cody 1986; Gentry 1986). Sin embargo,
esta distribución y baja densidad no sólo estaría
influenciada por parámetros biológicos y/o biofísicos
como el clima, si no también por agentes extrínsecos,
tales como los altos índices de deforestación que esta
región ha sufrido. De mantenerse esta tendencia, las
poblaciones de Ecuadendron
no podrán permanecer estables por generaciones y estarían
condenadas a la extinción. Estas características, es
decir, tamaño poblacional pequeño y distribución
geográfica restringida, agravadas por la pérdida del
hábitat, ponen a Ecuadendron
en un estado crítico de extinción, por lo que las
Reservas Ecológicas en las que se encuentran algunas de las
poblaciones de Ecuadendron
ofrecen una gran oportunidad para implementar programas para la
conservación in
situ
de esta leguminosa endémica del Ecuador.
Agradecimientos
Agradezco
a la Elizabeth Bascom Fellowship y al Missouri Botanical Garden sin
cuyo financiamiento y apoyo esta investigación no habría
sido posible. La fase de campo fue realizada con fondos de la Baltzer
Fellowship y del International Center for Tropical Ecology (ICTE) de
la University of Missouri-Saint Louis. Un especial agradecimiento a
Bette Loiselle, Kimberlie McCue y David Neill por sus comentarios y
discusiones durante la preparación de los borradores de este
trabajo. De la misma manera, agradezco a la especialista en SIG,
Trisha Consiglio, del Missouri Botanical Garden, por su significativo
aporte en las aplicaciones de las diferentes versiones de ArcView®.
Rodrigo Sierra de la University of Texas, generosamente facilitó
los mapas de uso de tierra y cobertura vegetal. Agradezco al
Ministerio del Ambiente del Ecuador, especialmente a la Blga. Mireya
Pozo, y a los funcionarios y guardaparques de la Reserva Ecológica
Manglares-Churute por el soporte logístico. Así mismo,
agradezco a Freddy Villao por la asistencia de campo, y a Digna
Palacios, Rodrigo Prado y Bolívar García, por
permitirme el acceso a sus propiedades en el área de Manta
Real. Cabe resaltar el apoyo de los miembros del Jardín
Botánico de Guayaquil, especialmente de Arcadio Arosemena y
Marcelo Bejarano. Finalmente agradezco los comentarios y
observaciones de mis colegas Carlos Daniel Cadena, Renata Durães,
José Fabara, Patricia Feria, Kimberly Holbrook, Andrea
Loaiza, Fátima Piña-Rodriguez, Seema Shet y
Wendy Tori.
Referencias
Ågren,
J. 1996. Population size, pollinator limitation, and seed set in the
self incompatible herb Lythrum salicaria. Ecology
77 (6): 1779-1790.
Blackburn,
T. M. & K. J. Gaston. 1997. Who is rare? Artifacts and
complexities of rarity determination. Pp. 48-69 en: W. E. Kunin
& K. J. Gaston (eds.). The
biology of rarity. Chapman and
Hall. London.
Carpenter,
G.; A. N. Gillison & J. Winter. 1993. DOMAIN:
a flexible modeling procedure for mapping distribution of plants and
animals.
Carulli,
J. P. & D. E. Furbrothers. 1988. Allozyme variation in three
eastern US species of Aeachynomene (Fabaceae), including the
rare A. virginica. Systematic
Botany13: 559-566.
Cody,
M. 1986. Diversity, rarity and conservation in Mediterranean-climate
regions. Pp. 122-152 en: M. E. Soulé (ed.).
Conservation Biology: The science
of scarcity and diversity. Sinauer
Assoc., Sunderland, MA.
Factos,
M. 2000. Alternativas
de manejo para la conservación del Bosque Protector
Molleturo-Mullopungo Comunidad Manta Real.
Fundación Natura/PNUD/Comunidad de Manta Real.
Feinsinger,
P.; H. M. Tiebout & B. E. Young. 1991. Do tropical
bird-pollinated plants exhibit density-dependent interactions? Field
experiments. Ecology
(72): 1953-1963.
Foster,
R. B. 1992. Biogeographic overviews. Vegetation.
Pp. 58 en: T. A.
Parker
III & J. L. Carr (eds.). Status
of the forest remnants in the Cordillera de la Costa and adjacent
areas of southwestern Ecuador.
Conservation International. RAP Working Papers 2.
Foster,
R.; N. Hernández; E. Kakudidi & R. Burnham. 1998.
Rapid assessment of tropical plant
communities using variable transects: an informal and practical
guide (ms. unpublished).
Gaston,
K. J. 1997. What is rarity? Pp. 30-47 en: W. E. Kunin & K.
J. Gaston (eds.).The Biology of
rarity. Chapman and Hall. London.
Gentry, A. H. 1986. Endemism in tropical
versus temperate plant communities. Pp. 153-181
en: M. E. Soulé (ed.). Conservation
Biology: The science of scarcity and diversity.
Sinauer Assoc., Sunderland, MA.
Goodman,
D. 1987. The demography of chance extinction. Pp. 11-34 en: E.
M. Soulé (ed.). Viable
populations for conservation.
Cambridge University Press. Cambridge.
Hendrix, S. D. & J. F. Kyhl. 2000.
Population size and reproduction in Phlox pilosa.
Conservation Biology
14 (1): 304-313.
Herendeen,
P. S.; A. Bruneau & G. P. Lewis. 2003. Phylogenetic relationships
in thecaesalpinioid legumes: a preliminary analysis based on
morphological and molecular data. In: B. B. Klitgaard and A. Bruneau
(editors). Advances in Legume
Systematics, part 10, Higher Level Systematics.
Pp. 37'62. Royal Botanic Gardens, Kew.
Hijmans,
R. J.; S. Cameron & J. Parral. 2004. Worldclim,
version 1.2. A square kilometer resolution database of global
terrestrial surface climate.
Available at http://biogeo.berkeley.edu/
INEFAN
- FUNDACI-N NATURA/ECOLAP - USFQ, 1999. Plan
de manejo de la Reserva Ecológica Manglares-Churute.
Divaprint. Quito.
Kunin,
W. E. 1993. Sex and the single mustard: population density and
pollinator behavior effects on seed-set. Ecology
(74): 2145-2160.
Kunin,
W. E. 1997. Population size and density effects in pollination:
pollinator foraging and plant reproductive success in experimental
arrays of Brassica kaber. Journal
of Ecology 85: 225-234.
Lande,
R. 1988. Genetics and Demography in Biological Conservation.
Science
241: 1455-1459.
Loiselle, B. A.; C. A. Howell; C. H.
Graham; J. M. Goerck; T. Brooks et al. 2003. Avoiding pitfalls of
using species distribution models in conservation planning.
Conservation Biology
17 (6): 1591-1600.
MacArthur,
R. H. & E. O. Wilson. 1967. The
theory of island biogeography.
Princeton Univ. Press, Princeton.
Main, A. R. 1981. Rare species: Precious or
dross? Pp. 163'174 en: R. H. Groves & W. D. L. Ride (eds.).
Species at risk. Research in
Australia. Springer-Verlag Berlin
Heidelberg New York.
Matthies,
D.; I. Bräuer; W. Maibom & T. Tscharntke. 2004. Population
size and the risk of local extinction: empirical evidence from rare
plants. Oikos
105: 481-488.
McKinney, M. L. 1997. Extinction
vulnerability and selectivity: combining ecological and
paleontological views. Annual
Reviews of Ecology and Systematics
28: 495-516. Annual Reviews Inc.
Mejía,
L. E.; J. Acosta & P. de la Torre. 1990. Estudio
de cambios en el uso del suelo.
Fundación Natura. Quito.
Menges,
E. S. 1991. Seed germination percentage increases with population
size in a fragmented prairie species. [[i+]Conservation
Biology5: 158-164.
Mittermeier,
R. A.; N. Myers & C. G. Mittermeier. 1999. Hotspots.
Earth's Biologically Richest and Most Endangered Terrestrial
Ecoregions. Conservation
International.
Myers,
N.; R. A. Mittermeier; C. G. Mittermeier; G. A. B. da Fonseca &
J. Kent. 2000. Biodiversity hotspots for conservation priorities.
Nature
403: 853-858.
Neill, D. A. 1998.
Ecuadendron
(Fabaceae: Caesalpinioideae: Detarieae): A new arborescent genus from
Western Ecuador. Novon
8: 45-49.
Nitecki, M. H. (ed.). 1984. Extinctions.
University of Chicago Press. Chicago.
Peña-Claros, M. 2003. Changes in forest structure and species
composition during secondary forest succession in the Bolivian
amazon. Biotropica
35(4): 450-461.
Pitman, N.C.; A. J.
Terborgh; M. R. Silman & P. Nuñez. 1999. Tree species
distribution in an upper amazonian forest. Ecology
80(8): 2651-2661.
Reveal, J. L. 1981. The concepts of rarity
and population threats in plant communities. Pp.
41-47 en: L. E. Morse & M. S. Henifin (eds.). Rare
plant conservation: Geographical data organization.
The New York Botanical Garden. New York.
Shaffer,
M. L. & F. B. Samson. 1985. Population size and extinction: a
note on determining critical population sizes. The
American Naturalist 125(1):
144-152.
Sierra, R. (ed.). 1999.
Propuesta preliminar de un Sistema
de Clasificación de Vegetación para el Ecuador
Continental. Proyecto
INEFAN-GEF-BIRF y EcoCiencia. Quito, Ecuador.
Soulé,
M. E. (ed.). 1987. Viable
populations for conservation.
Cambridge
University
Press. Cambridge.
Terborgh,
J. & B. Winter. 1980. Some causes of extinction. Pp. 119-133
en: M. E. Soulé& B. Wilcox (eds.). Conservation
Biology: an ecological-evolutionary perspective.
Sinauer Assoc. Sunderlan, MA.
Valencia, R.; N. Pitman; S. León-Yánez
& P. M. Jørgensen (eds.). 2000.
Libro rojo de las plantas endémicas
del Ecuador 2000. Herbario QCA,
Pontificia Universidad Católica del Ecuador, Quito.
Watkinson,
A. R. & J. L. Harper. 1978. The demography of a sand dune annual:
Vulpia fasciculata. I. The Natural regulation of populations.
Journal of Ecology
66: 15-33.
Zhou, G. 2001. Area
calculation for polygons.
http://gis.esri.com/arcscript.cfm.
Table 1. Gradiente altitudinal de la ubicación
de los transectos variables en los sitios de estudio Churute 1,
Churute 2 y Manta Real.
Area Nombre del transecto
CHURUTE 1
|
Número del
transecto
|
Gradiente
altitudinal (m)
|
Cerro
Masvale
Masvale
|
1
|
20
- 640
|
|
Cerro
Pancho Diablo
|
|
|
Cumbre 1
Cumbre 2
Mango
|
3
|
20
- 727
|
|
CHURUTE 2
|
|
|
Cerro Perequeté
P-Grande
|
1
|
20
- 408
|
Cerro Simalón
Carrillo-Toro
Maridueña
|
2
|
20
- 668
|
Cerro El Mate
Mate
|
1
|
20
- 337
|
MANTA REAL
|
|
|
B-García
|
1
|
300 - 625
|
S-Prado
|
1
|
360 - 950
|
La Aurora
|
1
|
400 - 480
|
R-Prado
|
1
|
520
- 650
|
Caimatán
|
1
|
560 - 630
|
Paquisha-La Victoria
|
1
|
520 - 900
|
Sanaguín
|
1
|
600 - 1105
|