Introducción

Proyectos para el
estudio y la descripción de formaciones de vegetación
tropical están muy complicados por la inmensa diversidad de
especies de la región, mosaicos de vegetación muy
complejos y problemas de taxonomía y logística.
Inventarios detallados requieren estudios a largo tiempo para lograr
incorporar todas los especies. Estudios florísticos para la
descripción de formaciones de vegetación tropical se
complican inmensamente por la increíble diversidad de especies
de la región, su mosaico de tipos de vegetación muy
complejo, problemas taxonómicas, logísticas y escasez
de tiempo disponible. Estudios detallados requieren trabajo de campo
a largo tiempo por causa de la multitud de especies encontrados en
estado infértil durante visitas cortas. Por estas razones el
número de estudios de vegetación esta muy limitado
particularmente en los Neotropicos, y solo de pocos lugares se conoce
inventarios florísticos mas o menos completos. Los pendientes
extremos especialmente en el área andina causa una diversidad
de habitatas sorprendente que permite distinguir entre pequeños,
homogéneos tipos de vegetación forestal.
El problema mayor
para estudios fitosociológicos en bosques húmedos de la
zona baja se presenta por la densidad muy baja de las poblaciones de
muchos especies, causando dificultades en establecer las respectivas
áreas-mínima. Este problema no se presenta en los
bosques montanos. El terreno sumamente escarpado causa una gran
diversidad de habitats que permite distinguir entre pequeños y
homogéneos asociaciones de bosques.
Hasta recién,
estos bosques montanos tropicales - aunque, sumamente
importante en manera ecológica y económica como
captaciones de agua y protección contra erosión -
recibieron solo una atención marginal de ciencia y sociedad.
Peor, la mayoría de los estudios hechos en los ecosistemas de
las montañas tropicales se concentraron en la zona alpina,
mientras los bosques, con frecuencia casi inaccesibles, con su imensa
diversidad de especies casi no fueron estudiados. Aún
proyectos grandes como "Ecoandes" en Colombia (Hammen et
al. 1983, 1984, 1989, 1995) incluyeron la región de los
bosques solo en manera marginal. Las pocas publicaciones sobre la
vegetación de los bosques montanos de Ecuador solo contienen
listas de especies o mencionan la región montana en
comparación de los bosques de la Amazonía (Grubb et al.
1963, 1966; Ek 1997). Los primeros esfuerzos de unir este
conocimiento inicial se hicieron recién (Hamilton et al. 1994,
Churchill et al. 1995). El déficit de información
científica especialmente incluye los aspectos de regeneración
y uso de los bosques montanos y los procesos de sucesión
después de impactos naturales o antrópicos. Los
requisitos del hábitat y el potencial para regeneración
de los especies maderables importantes están casi
completamente desconocidas.
La
región fronteriza de Ecuador y Perú (Fig. 1) pertenece
a los áreas biológicamente más diversas del
mundo y por este es un "punto caliente de biodiversidad"
por excelencia. Puertos bajos en la cadena andina permiten el
intercambio fácil entre las floras y faunas de la Amazonía
y la zona baja del Pacifico. Adicionalmente la región muestra
una transición muy rápida entre los Andes húmedos
del Norte y los bosques secos, deciduos de la zona baja del Norte de
Perú. Hasta recién, Parque Nacional Podocarpus y el
área de estudio estuvieron científicamente casi
desconocidos. Pocos estudios existen sobre la flora de la Provincia
de Loja (Espinosa 1948a,b; Emperaire & Friedberg 1990, Øllgaard
& Madsen 1993, Ulloa & Jørgensen 1993, Jørgensen
& Ulloa 1994, Madsen & Øllgaard 1994, Bussmann &
Lange 1998, Jørgensen & León-Yanez 1999), o tratan
descripciones cortas de su vegetación (Espinosa 1989/92,
Madsen 1989, 1991, Jørgensen 1991, Bøgh 1992). El
esfuerzo mas nuevo para la clasificación de la vegetación
de Ecuador (Sierra 1999) clasifica todos los bosques montanos entre
1800 - 3000 m de altitud como "bosque de neblina
montano" sin mas distinción.
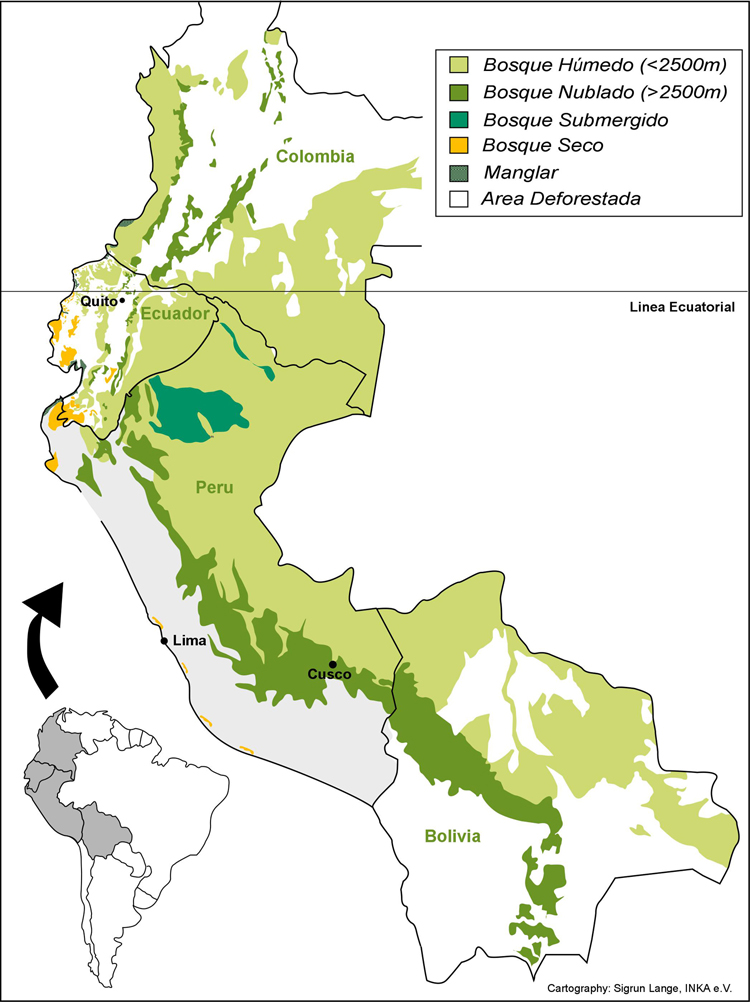
Fig. 1: La Region Andina y el enfoque del estudio.
Estudios
sobre la composición y regeneración de los bosques de
Reserva Biológica San Francisco se elaboraron desde 1997 en el
margen del proyecto "Funcionalidad en un bosque montano
tropical: Diversidad, procesos dinámicos y
potencial para el uso" de la Fundación Alemana para la
Investigación (DFG). La intención de las
investigaciones fue de caracterizar los diferentes tipos de bosques y
sus estados de regeneración a lo largo de la gradiente
altitudinal, y de evaluar si una caracterización detallada se
puede elaborar solo a base de un estudio fisiognómico o si se
necesita una análisis fitosociológico detallado. Desde
2001, se desarrollaron estudios sobre el uso de plantas medicinales
en el Norte de Perú, dentro del programa de investigación
y entrenamiento para estudiantes de minorías (MIRT) del
instituto Nacional de Salud (NIH).
Reserva
Biológica San Francisco se encuentra entre las capitales
provinciales Loja y Zamora e incluye 1000 hectáreas de la
extensión norte de la Cordillera de Consuelo, a 03°58'18''S
- 079°04'44''W, en la provincia
Zamora-Chinchipe, Ecuador, bordeado por el Parque Nacional
Podocarpus, la única área protegida en el sur del país.
Con extensión de 1800 - 3150 m la ECSF incorpora un
transecto completo de los bosques montanos de la región y
áreas antropicamente perturbadas para comparación (Fig.
2). La topografía es inmensamente escarpada con pendientes
entre 40-60° y frecuentemente llegando a 90°.
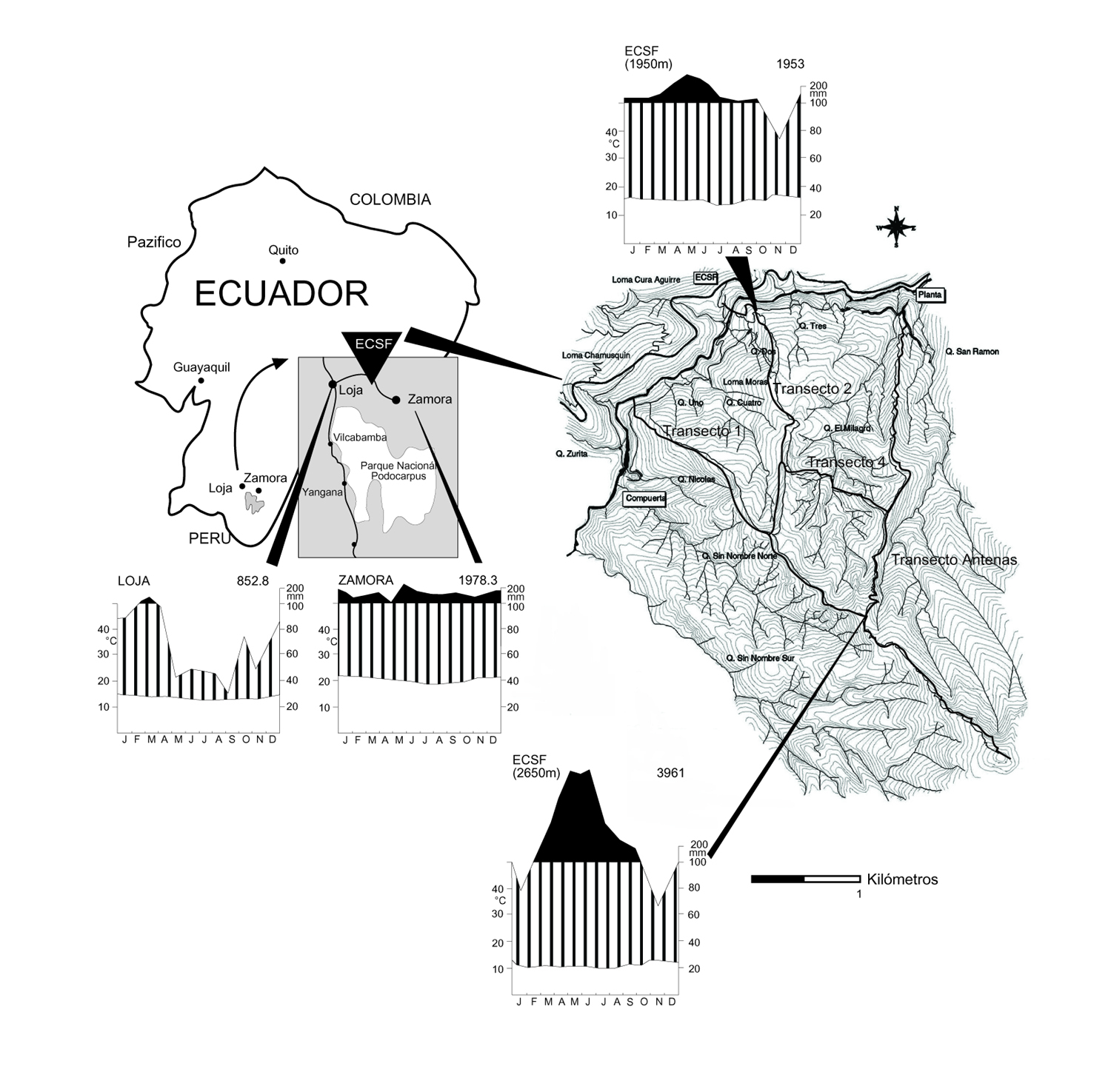
Fig. 2: El terreno de la Estacion Científica
San Francisco, Ecuador.
En
la parte Sur de Ecuador y el Norte de Perú la elevación
mas alta es de 4600 m, y por gran parte no se supera 4000 m
(Jørgensen & Ulloa Ulloa 1994). Esta área
representa la región mas baja en los Andes ecuatoriales.
Mientras la piedra base del Andes del Norte esta formado por
volcanismo Quarterno, la parte Sur esta formado por sedimentos
Terciacos (Hall 1977). Los sustratos geológicos consisten de
roca arenosa y piedra azul, los suelos son por gran parte
Dystrudepts, Humaquepts y Petraquepts (Schrumpf 1999).
Emck (in prep.)
describe una precipitación anual promedia de 2500 mm en la
zona baja, y mas que 10000 mm en las zonas mas altas de esta parte de
la Cordillera. Especialmente la parte mas alta se encuentra casi en
nubes o neblina todo el año. La precipitación mayor se
encuentra en Febrero-Marzo y Junio-Setiembre, ambos periodos seguidos
por épocas mas secas que pueden causar cortas temporadas casi
áridas en particular en los meses Octubre-Enero.
Materiales y Métodos
Después
de un estudio florístico detallado, los bosques de la Reserva
Biológica San Francisco (ECSF) han sido investigado en
estudios fisiognómicos y fitosociológicos. Mas que
trecientocinquenta parcelas en áreas de bosques homogéneas,
120 parcelas en derrumbes naturales y 150 en derrumbes antropogénicas
fueron investigadas con la metodología de Braun-Blanquet
(1964) descrito por Mueller-Dombois & Ellenberg (1974) y
modificado por Hammen et al. (1989). El tamaño de las parcelas
(mínimo 400 m2 en bosques y 4 m2 en
caso de derrumbes) siempre estuvo mas grande que el área
mínima determinada (2m2 para derrumbes, 256m2
para el bosque montano bajo y 64m2 para el bosque montano
alto y áreas de Jalca.
Para una analices
del uso de bosque se hizo entrevistas con curanderos, pobladores y
carpinteros en las provincias de Loja y Zamora-Chinchipe en Ecuador,
y con curanderos en La Libertad y Lambayeque en el Norte de Perú.
Las muestras
colectadas fueron identificados y depositados en los herbarios TRUX,
HUT, LOJA, ECSF, QCNE y QCA.
Resultados
Zonación
de la vegetación en los Bosques Montanos del Sur de Ecuador y
Norte de Perú
Taludes muy
escarpados y quebradas muy profundas causan un mosaico de diferentes
condiciones micro climáticas, que, junto con la frecuencia de
derrumbes naturales causa un mosaico muy variable de unidades de
vegetación. Basado en los datos florísticos los bosques
montanos de la ECSF se agruparon en tres formaciones mayores, cada
una ocupando una zona de altitud especifica (Bussmann 1999, Bussmann
2001.).
Bosque
Montano Bajo
La
formación mas importante de bosque entre 1200 - 2100 m,
con un dosel muy diverso de 2 estratos esta el "Bosque montano
bajo" (bosque de Ocotea - Nectandra),
extendiendo hasta 2300 m al fondo de quebradas con protección
contra el viento. Especies de familias perteneciendo al bosque húmedo
tropical (Cyclanthaceae, Lauraceae, Piperaceae) se encuentran muy
común en estos bosques, mientras que especies de la flora de
la zona alta están raras. áreas primarias del bosque de
Ocotea-Nectandra se encuentran especialmente en filos muy
escarpados con inclinaciones de 30-50° o mas, y en quebradas casi
inaccesibles. En regiones de fácil acceso esta formación
ha sido casi destruido por actividades antropicas y reemplazado por
bosques secundarios.
Con 880-1210 árboles
de mas de 10cm diámetro al altura del pecho (dap)/ha, el
Bosque Montano Alto muestra la abundancia mas alta de árboles
en el área de estudios, y una de la abundancias más
altas encontrado en bosques montanos hasta ahora. Con 339 especies
las epifitas son la forma de vida mas diversa en estos bosques,
seguido por árboles (318 especies), hierbas (186), arbustos
(134), helechos no-epifititos (108) y trepadoras (26) y parásitos
(4) (Fig. 3). La mitad de los géneros de Araceae encontrados
en ECSF crecieron solo en esta zona baja. Orquídeas son el
grupo de epifitas dominante (153 especies), con casi la mitad de los
géneros restringidos a esta región.
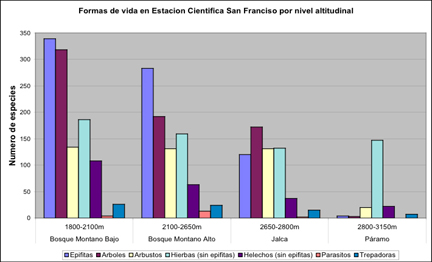
Fig. 3: Distribución de formas de vida en
diferentes niveles de altitud.
-
Donde el bosque clímax ha sido destruido - probablemente
por fuegos naturales - el estrato arbóreo esta dominado
por palmas grandes (Arecaceae, Dictyocaryum lamarckianum). El
estrato de hierbas esta dominado por diferentes bambusaceas (Chusquea
spp.), y Gleicheniaceae (Dicranopteris spp., Sticherus spp.)
casi impenetrables. La regeneración de los especies del bosque
primario parece inhibido en estos áreas.
-
En lugares con mucho impacto antropico (tala y quema), se desarrolla
un bosque secundario monotípico, completamente diferente. Esta
comunidad solo tiene un solo estrato arbóreo, dominado
completamente por los fustes de Axinea quitensis
(Melastomataceae), con una altura de 10-12 m. La diversidad de
especies esta mucho mas baja, y aparte de Axinea, solamente
Vismia tomentosa (Clusiaceae), otra especie pionera contribuye
al dosel con mayor número de individuos. Casi no se encuentran
epifitas, y la flora herbácea también esta muy
empobrecida. No existe datos para clarificar si esta comunidad
secundaria esta estable, o si se pueda desarrollar a una vegetación
primaria después de cierto tiempo.
Bosque
Montano Alto
A
partir de altitudes de mas que 2100 m, hasta mas o menos 2750 m, el
"Bosque montano alto" (bosque de Purdiaea nutans -
Myrica pubescens - Myrsine andina), una formación
monotípica con un solo estrato arbóreo de una altura de
5-10 m, a veces llegando hasta 15 m, reemplace el bosque de
Ocotea-Nectandra. Especies de la zona baja casi desaparecen
por completo. El dosel esta completamente dominado por los fustes
torcidas de Purdiaea nutans (Cyrillaceae), que tiene su mayor
distribución en el Norte de Perú. Existe un estrato muy
diverso de arbustos y árboles pequeños. Sobre 2450 m de
altitud, especialmente en filos expuestos al viento se encuentra la
transición al "bosque subalpino", y a mas que 2650
m ya existen islas de "Páramo". Derrumbes muy
frecuentes mantienen los procesos dinámicos en el bosque
montano alto y son muy importantes para el mantenimiento de la
diversidad. En bosques muy viejos se diminúye la diversidad
florística rápidamente.
La posición
sintaxonomica del Bosque Montano Alto todavía requiere
clarificaciones. Este orden nuevo muestra relaciones a la "grupo
de comunidades de Hedyosmum pseudoandromeda" establecido por
Meier (1998).
El número de
árboles se diminuye claramente. A 2225 m, se encuentran 650
arboles/ha todavía, bajando a 160 en 2425 m, donde el bosque
tienen una apareciencia muy abierta. Epifitas todavía son la
forma de vida mas abundante con 283 especies. La diversidad de
árboles y pteridofitos no-epifititos se diminuya rápidamente
(192 y 63 especies), hierbas (159) y arbustos (131) se vuelven más
importante que en el bosque Montano Bajo, como en la misma manera
trepadoras (24) y parásitos (13) (Fig. 3). Aunque la mayoría
de las familias importante se mantiene, su número de especies
decrece, con orquídeas como excepción notable.
Especialmente Pleurothallidinae (Lepanthes, 32 especies;
Pleurothallis 25 especies) tienen su centro de distribución en
estos bosques de neblina. Polypodiaceae epifititas pequeñas
como Terpsichore y Melpomene se encuentran con mayor número de
especies en esta altitud, y Hymenophyllaceae también tienen su
centro aquí.
-
En claros naturales, especialmente en áreas planas que
probablemente están influidas por el efecto de fuegos
naturales, el estrato arbóreo casi desaparece, con muy pocos
ejemplares viejos de árboles permaneciéndos,
dejando espacio para un estrato muy denso de la Bambosoidea
Neurolepis elata, suprimiendo casi todos los otros especies
del estrato herboreo. La floración masiva, seguido por el
muerte de Neurolepis se han observado 2001. Si este vaya a
causar un cambio de vegetación no esta claro hasta ahora.
-
En años muy secos, parece que se desarrollan fuegos también
en áreas normalmente muy húmedas. La intensidad de
estos fuegos parece mucho mas bajo que en los áreas secas, y
por eso solo los estratos arbustivos y herbáceos están
destruidos, mientras el dosel se mantiene. El piso del bosque esta
frecuentemente cubierto por Sphagnum sp. En vez de Líquenes.
Cyperaceae grandes (e.g. Rhynchospora locuples) están
formando una alfombra graminosa.
Jalca
(Bosque Subalpino, Ceja Andina)
La
zona de bosque mas alta esta formada por el "bosque subalpino",
que tiene una apareciencia similar a la "Jalca" de
Bolivia. Esta formación, conforma una vegetación
arbustiva casi impenetrable que un bosque esta densamente
interrelacionada con los Páramos. El limite de bosque en el
área de ECSF esta causado mayoramente por los vientos muy
fuertes. áreas con protección contra el viento se
encuentran densamente cubiertas por Jalca, mientras áreas con
exposición al viento están ocupados por Páramo a
la misma altitud. Por eso, no existe un limite de bosque real en la
región. A partir de una altitud de 2450 m se puede encontrar
islas de Jalca ya en el bosque montano alto. Las copas de las
especies dominantes - especialmente Cunoniaceae (Weinmannia
spp.), Clusiaceae (Clusia spp.), Clethraceae (Clethra
spp.) y muchas Melastomataceae pequeñas (Brachyotum
spp., Miconia spp.) solo tienen 1-2 m de diámetro,
formando un dosel sumamente denso, con muy poca luz llegando al piso.
Los fustes dan origen a una alfombra muy profunda de briofitos y
material orgánica, y en diferencia a la diversidad de la flora
arbustiva, la flora herbácea esta muy pobre.
Árboles
representan la forma de vida mas abundante con 172 especies, arbustos
(131), hierbas (132), helechos no-epifitos (37), trepadoras (15) y
parásitos (2). Solo se registraron 120 especies de epifitas en
la Jalca. Familias de la zona baja como Araceae y Piperaceae
desaparecieron, la diversidad de Bromeliaceas y Orquídeas ha
disminuido. Cuando en otros tipos de bosque las Orquídeas
representaron más o menos una tercera de las epifitas, en la
Jalca su importancia llega a 60 %. Este esta muy comparable con los
resultados de Bgh (1992) que encontró 138 especies en
una parcela en el área cercana de Cajanuma. Las comunidades de
Jalca crecen sobre Oxaquic y Aquic Dystropepts y reciben una
precipitación anual de casi 4000 mm. La ocurrencia casi
constante de neblina les da humedad adicional.
Páramo.
Páramo
Los
Páramos de la región son sumamente ricos de especies, y
con frecuencia cubiertos por nubes, con la precipitación anual
llegando a 10000 mm. La frecuencia de vientos fuertes (con un
promedio de velocidad máxima del viento de 60 km/h) esta
característica por el área. Typic Tropaquepts y Lithic
Troporthents son los tipos de suelos prevalentes. Las gramíneas
mayores son Bambusoidas del género Neurolepis. Estos especies
son obviamente muy sensitivos al efecto de fuegos frecuentes, y por
eso están reemplazados de inmediato por gramíneas más
duras como Calamagrostis, Festuca y Stipa en
condiciones con quemas frecuentes. Por la razón que estos
géneros no tienen mucha importancia en el Neurolepion típico
se puede clasificar los Páramos de la ECSF como muy naturales.
Con su abundancia muy alta de diferentes especies de Neurolepis,
los Páramos del área de estudio pueden ser
interpretados como remanentes de la vegetación alpina natural
potencial que probablemente ha cubierto áreas grandes de la
montaña alta del Sur de Ecuador originalmente, mientras los
Páramos de gramíneas duras se encuentran en regiones
perturbadas por impacto antropico y deben ser considerados como
vegetación secundaria. Árboles (3 especies), Epifitas
(4) y trepadoras (7) casi desaparecen. Arbustos (20) y helechos
no-epifiticos (22) se vuelven menos importantes, mientras hierbas
(147 especies) son la forma de vida mas importante de los Páramos
(Fig. 3). Sintaxonomicamente los Neurolepio-Puyetalia están
parte de los Wernerietea Cleef 1981.
Ecología
de regeneración de bosques montanos tropicales
Los Bosques Montanos
de América Latina, de cuales solo pocos se quedan después
de tala, muestran ciclos típicos de regeneración
mosaica. Bajo condiciones naturales los claros que son resultado de
caída natural de árboles, se cierran por especies que
llegan primero. En muchos casos no obstante se puede observar un
juego muy típico de especies pioneras de crecimiento rápido,
que actúan como árboles de sombra, bajo este dosel se
desarrollan las especies clímax, que necesitan sombra para su
germinación y crecimiento juvenil. Esta regla, que sirve para
muchos bosques tropicales, no obstante, muestra problemas en ciertos
bosques, los cuales tienen una especie dominante o especialmente
importante. Uno de los mejores ejemplos de este se encuentra en
África:
En
el Bosque Montano Bajo del Sur de Ecuador y Norte de Perú, los
claros están colonizados de inmediato por Cecropiaceae
(especialmente Cecropia montana), Piptocoma discolor
(Asteraceae), Isertia laevis (Rubiaceae) y Heliocarpus
americanus (Tiliaceae). El dosel se cierre muy rápido, y
los especies primarias se regeneran bajo la sombra de los pioneras.
Regeneración
de derrumbes naturales en el ambiente Andino
En
los Andes Ecuatorianos, los derrumbes fueron estudiados especialmente
por Stern (1995) y Kessler (1999) quien trabajo en Bolivia. Otros
trabajos sobre derrumbes en el ambiente tropical incluyen Garwood et.
al (1979) en Panamá, y Guariguata (1990) en Puerto Rico. En
estas regiones tropicales, derrumbes se encuentran con gran
frecuencia. En adición a pendientes escarpados y lluvias
fuertes, la construcción de carreteras y tala y quema
debilitan el substrato, y la descomposición del material
geológico también tiene una influencia en la frecuencia
de derrumbes (Brabb & Harrold, 1989).
La función de
derrumbes para el desarrollo de la vegetación de esta región
es muy importante, y representan un factor sumamente importante por
el mantenimiento de la biodiversidad, la dinámica y la
estabilidad del bosque montano. En bosques muy viejos, especialmente
en el Bosque Montano Alto, la diversidad esta drásticamente
reducida. La mayoría de las especies encontrados durante el
proceso de la sucesión no son elementos del bosque maduro. Por
eso, los derrumbes son un factor muy importante para la regeneración
del ecosistema.
La
sucesión de derrumbes en Ecuador sigue mas o menos el esquema
encontrado en otras áreas de montaña (Herzog 1923,
Simonett 1967, Garwood et al. 1979, Guariguata 1990,
Stern 1995). No obstante, como Kessler (1999) ya observó
en Bolivia, la regeneración de derrumbes Andinos muestra
diferencias en relación a la función de diferentes
helechos. Como en Bolivia, Pteridium arachnoideum -
aunque muy importante en la sucesión de áreas
antropicamente perturbadas - no tuvo ninguna importancia en la
regeneración de derrumbes naturales en el Sur de Ecuador, con
Gleicheniaceae encontrados como los helechos mayores. A diferencia
muy clara a Stern (1995) en el Norte de Ecuador y Kessler
(1999), Bambú (Chusquea spp.) no tuvo ninguna
importancia en la regeneración de derrumbes en la Estación
Científica San Francisco. Gramíneas como Cortaderia
tampoco tuvieron una influencia larga en los proceso de sucesión.
Por el contrario a los observaciones de Kessler (1999), casi
no se observo bosques muy viejos dominados por helechos o Bambú.
Gramíneas como Neurolepis elata o Rhynchospora
locuples se encontraron gran parte donde la comunidad terminál
hubiera sido destruido, probablemente por fuegos naturales o estuvo
muy viejo. Se debe iniciar investigaciones de los procesos de
sucesión a largo tiempo en todo de la región andina
para salir con datos generales sobre la regeneración.
En
situaciones naturales, la primera fase de sucesión de
derrumbes esta dominada por briofitos, especialmente Polytrichum,
y un estrato diverso de líquenes. Después de un tiempo
entran especies de plantas vasculares. La duración de la
regeneración depende mucho en la intensidad de actividad en el
derrumbe. En promedio, la fase de criptógamas dura mas o manos
cinco años. En localidades con suelo muy rocoso la erosión
puede continuar por mucho tiempo. En esta situación,
representada con frecuencia al lado de carreteras, la fase primaria
se puede extender por mucho tiempo.
La
segunda fase de sucesión muestra un incremento de la densidad
vegetal. Individuos de Gleicheniaceae y Lycopodiaceae colonizan los
derrumbes. Lycopodiella glaucescens y Lycopodium clavatum
crecen mas rápido que las Gleicheniaceas, pero forman colonias
menos densas. Esta fase secundaria esta mayoramente caracterizada por
competencia con los pioneros de la primera fase, y por parte ya
empieza regeneración de especies arbustivas como Tibouchina
lepidota, Graffenrieda harlingii (Melastomataceae) o Bejaria
aestuans (Ericaceae). Estas pioneras no obstante no se encuentran
en la comunidad clímax. En una fase tercera, la vegetación
esta dominada por arbustos como Ageratina dendroides
(Asteraceae), y al fin se encuentran especies del bosque secundario.
Regeneración
de bosques monotípicos y Jalca
En
los bosques monotípicos como el Bosque Montano Alto en
Ecuador, los ciclos de regeneración después de eventos
de fuego, tienen mayor importancia.
En
áreas demasiada viejas del Bosque Montano Alto y Jalca en el
Sur de Ecuador, el piso del bosque se encuentra densamente cubierto
por Neurolepis elata (Bambusoideae), y el dosel parece muy
abierto y diminuido, con cubierta del dosel de menos que 5%. Eventos
de floración masiva de Neurolepis, y de la misma menara
eventos de sequía, los cuales pueden ocurrir durante periodos
de El Niño / La Niña, se presenta un monto elevado de
material orgánico seco, y por eso una situación
excelente por fuegos. Después de la quema, los áreas
abiertas se colonizan rápidamente por especies de helechos,
sin un estado criptógamo. En un tiempo muy corto se establecen
especies arbóreos, en particular Purdiaea nutans,
llegando de los bosques primarios de los alrededores, y después
de un par de años las Bromeliáceas características
Hacen su arribo. Debido al crecimiento muy lento de las especies
dominantes, se desarrolla un estrato arbustivo muy denso, que se
puede mantener por muchos años. Purdiaea nutans empieza
de sobrepasar el resto de la vegetación lentamente, formando
el típico clímax monotípico. Después de
mas o menos 4-500 años, este estrato denso de Purdiaea
empieza a morir, y el estrato denso de Bromeliáceas y arbustos
se cambia por las Bambusoideas mencionadas, tolerantes de mucha
insolación.
Uso de
Recursos Forestales
Estructura
del bosque y volumen de madera
841
individuos de árboles se marcaron en las parcelas,
representando 81 especies de 54 géneros y 38 familias. Los
géneros con mayor número de especies fueron Nectandra,
Miconia, Inga, Licaria, Clusia, Myricanthes y Panopsis.
Diversidad y número de especies se disminuyeron con la
altitud. Las familias mas importantes en el bosque montano bajo son
Lauraceae, Rubiaceae, Mimosaceae y Melastomataceae. En
la parte media de los transectos de diversidad y el número de
individuos se diminuyan, árboles con dbh mayor están
mucho mas raros a altitudes mayores. Las especies mas importantes en
siete niveles altitudinales no solo mostraron claramente que la
diversidad se disminuya con la altitud incrementando, si también
con el nivel de perturbación. A 1825 m, Pollalesta
discolor, especie pionera, se encontró mas común,
mientras las otras especies se encontraron solo esporádicamente.
Este indica un nivel alto de perturbación en esta región.
A altitudes de 1925, 2025 y 2125 m se encontró Graffenrieda
emarginata, Alzathea verticillata y Hedyosmum anisodorum como
especies mas abundantes. Finalmente a alturas mas altas, Purdiaea
nutans, Myrica pubescens y Clusia latipes monstraron el
número mas alto de individuos.
De los 81 especies
encontrados, diez fueron los mas importantes. No obstante solo en el
bosque montano alto se encontró una dominancia aparente de una
sola especie.
Extrapolado a una
hectárea, 880 fustes individuales se encontraron a 1850 m,
1180 a 1925 m, 1210 a 2025 m, 910 a 2125 m, 650 a 2225 m, 460 a 2325
m y 160 a 2425 m. (Tab. 1). En cada zona de altitud el tamaño
de la mayoría de los árboles estuvo muy pequeño,
con pocos individuos mas grandes (Bussmann et al. 2003).
El
área basal/m2 estuvo lo mas grande en el bosque
montano bajo, llegando a la altura de 2125 m, diminuyendo rápidamente
en zonas mas altas. El área basal de los especies
verdaderamente maderables sorprendentemente se mantuvo casi
constante, aunque el volumen de madera (VT) y el volumen de la madera
de los fustes (VC) también se disminuyeron con la altitud
incrementada (Tab.1).
Tab.
1. Datos sobre especies de árboles en Reserva Biológica
San Francisco
|
Altura
|
1825
m
|
1925
m
|
2025
m
|
2125
m
|
2225
m
|
2325
m
|
2425
m
|
|
Familias
|
20
|
17
|
22
|
14
|
13
|
10
|
8
|
|
Géneros
|
28
|
26
|
28
|
17
|
15
|
10
|
8
|
|
Especies
|
28
|
31
|
33
|
17
|
16
|
11
|
9
|
|
Individuos
/ ha
|
880
|
1180
|
1210
|
910
|
650
|
460
|
160
|
|
área
basal m2/ha
|
16,66
|
17,26
|
22,32
|
6,08
|
11,08
|
7,59
|
2,28
|
|
DAP
media
|
15,39
|
13,85
|
15,4
|
12,13
|
15,2
|
14,96
|
12,85
|
|
Altura
media
|
6,99
|
7,68
|
9,41
|
6,28
|
7,63
|
6,91
|
6,43
|
|
Altura
media del fuste
|
3,84
|
4,05
|
4,92
|
3,46
|
3,64
|
3,02
|
2,94
|
|
VC
m3/ha
|
67,12
|
54,45
|
87,58
|
20,38
|
31,12
|
17,88
|
9,37
|
|
VT
m3/ha
|
110,86
|
105,19
|
165,63
|
38,2
|
74,75
|
44,85
|
13,86
|
|
área
basal m2/ha (maderables)
|
4,85
|
7,16
|
7,54
|
4,15
|
4,01
|
7,17
|
1,59
|
|
VC
m3/ha (maderables)
|
20,6
|
26,98
|
31,81
|
15,01
|
11,96
|
16,56
|
3,95
|
|
VT
m3/ha (maderables)
|
35,47
|
47,24
|
66,64
|
28,23
|
25
|
46,95
|
10,06
|
La regeneración
natural mejor de todos los bosques se encontró a una altitud
de 1925 m, indicando que el bosque de esta altitud representa un
clímax-mosaico casi no perturbada. En los bosques de las
regiones mas altas la regeneración fue mas rara. Especialmente
en bosques viejos se encontró un sotobosque dominado por
gramíneas muy densas. Porque no existe perturbación
antropica en esta altitud se muestra claramente que estos bosques
necesitan perturbaciones naturales, por ejemplo fuegos, para permitir
la regeneración.
Forestería
y industria de madera en el Sur de Ecuador claramente reflejan la
crisis económica del país. La destrucción de los
hábitats en los bosques para su uso en agricultura y
ganadería, metodologías de tala no-sustentables y muy
poca información sobre posibilidades de reforestación
con especies nativas van a causar una escasez grave de madera en los
próximos años. Especies indígenas todavía
están en demanda muy alta, mientras las especies exóticas
como Eucalyptus globulus y Pinus radiata no tienen la
demanda de los usuarios. Como consecuencia la industria maderera,
incrementa la escasez de madera sin interés en la edad de
árboles, acelerando la crisis. La practica común de
tala - usando motosierras para cortar tablas directo en el
bosque, causa una perdida de la materia prima de hasta 50%, y por
eso, esta completamente no-sustentable. Por razón de la
destrucción amplia de los bosques para establecer pastizales,
tala selectiva , sin reforestación con especies nativas, una
escasez de madera ya es notoria en la industria maderera de la zona.
Cada vez más serio porque especies indígenas están
en demanda alta, y especies exóticas como Pinus radiata, P.
patula y Eucalyptus globulus no reciben mucho interés
del consumidor (Leischner & Bussmann 2003).
Para prevenir la
perdida de los bosques naturales, las autoridades ecuatorianas
pusieron restricciones para el uso de especies raras, tratando de
regular la tala. Desafortunadamente las regulaciones legales no están
muy conocidas en la población rural o no se obedecen. Un 50%
de toda la madera usada en el Sur de Ecuador esta de origen ilegal.
El control esta muy difícil y restringido por causa de escasez
de personal.
Continuando con la
practica presente, no solo de diminuirán los áreas de
bosques naturales, también la existencia de la industria
maderera misma se encuentra seriamente amenazada. Entendiendo el
problema, la población local misma tendrá interés
en reforestación con especies indígenas e inversión
en maquinaria mas avanzada para garantizar una eficaz y mas alta
producción del uso de los recursos naturales a largo plazo.
De
las especies maderables usadas al nivel local,
especialmente Cedro (Cedrela odorata L., Cedrela
montana Moutz ex Turcz, Cedrela lilloi C. DC),
Seique (Cedrellinga cateniformis Ducke),
Almendro (Swietenia macrophylla King),
Nogal (Juglans neotropica), Yumbingue (Terminalia
amazonia J. F. Gmel., Terminalia oblonga (Ruiz &
Pavón) Steudel) y Romerillo fino
(Podocarpus oleifolius Don ex Laub.), estan mas y mas
dificiles para encontrar, y arboles estan talados sin relación
a su edad o calidad. Plywood, MDF o metal estan usados mas y mas para
combatir la escasez de material. Como razones para la escasez
acelerando se menciona mayoramente variaciones climáticas,
restricciones nuevos del Ministerio de Medio Ambiente, competencia
incrementada y como razón mas importante "falta de
material prima".
Para combatir la
perdida de los bosques naturales el Ministerio de Medio Ambiente puso
restricciones para garantizar la cosecha controlada. Cada propuesta
de tala requiere un permiso escrito del Ministerio para cortar y en
la misma manera para el transporte de la madera. Desgraciadamente no
se conoce estas regulaciones en la población rural, y la tala
ilegal se encuentra con mucha frecuencia. El control policial
incrementa los precios de la madera y su escasez. Mas o menos 50% de
la madera en el mercado viene de la tala ilegal.
Mas que 80% de la
madera usada en Loja viene de los bosques naturales de Zamora. Solo
pocas especies estan taladas en los bosques del Sur de la provincia
de Loja. La mayoría de los productos de madera producidas en
Loja o Zamora estan vendidas dentro de las provincias. No obstante,
una parte considerable se vende a las provincias alrededores y a
Perú. Zamora solo tiene un papel muy pequeño en el
mercado interprovincial. La mayoría de los productos permanece
en la ciudad o se vende a Loja.
El
fenómeno climático de El Niño tiene cierta
influencia a las oscilaciones del mercado de madera, porque durante
temporadas con lluvias fuertes la única carretera entre Loja y
Zamora se encuentra bloqueada con frecuencia. Este problema no
obstante se mencionó solo en Loja, y no tiene influencia por
la disponibilidad de madera en Zamora. Ahí la migración
de una gran parte de mano de obra causa una escasez fuerte en
obreros, y en esta manera una disminución de producción.
En adición a
las actividades frecuentes de tala ilegal el procesamiento muy
ineficaz de la madera y las practicas no-sustentables de tala estan
los factores mas importantes para la destrucción de los
bosques de Ecuador. La mayoría de los arboles talados estan
cortados en tablas directamente en el bosque, usando motosierras.
Esto causa una perdida en forma de polvo de madera de hasta 50% del
material prima. La maquinaria muy vieja de la mayoría de
aserraderos y carpinterías - por razón de falta
de recursos económicos - incrementa la perdida del
material, que, hasta el producto final, puede alcanzar 75%.
Si el uso actual de
los bosques naturales continua, las condiciones rápidamente se
volvieran críticas para la industria maderera. Para evitar la
crisis se tiene que tomar medidas rápidas:
disminuir la
perdida de material durante tala y procesamiento de la madera
empezar con
programas de reforestación con especies indígenas
institucionalizar
programas de educación sobre uso sostenible de los bosques
Casi todas las
organizaciones prefieran la segunda opción. Por la razón
que la mayoría de las especies nativas crece comparativamente
lento, se necesita una acción rápida para instalar
sistemas agroforestales y silvopastoriles. No obstante, con este
escenario ideal la situación no se mejorará en los
próximos 10 años. Para mejorar el uso de la madera,
carpinterías y aserradotas, necesitan la posibilidad de
recibir créditos baratos para mejorar su maquinaría. El
establecimiento de un Parque Industrial en Loja seria el primer paso
en esta dirección.
A largo plazo, los
bosques naturales de la región son la parte mas importante con
que trabajar. Su protección no solo depende en los
agricultores y ganaderos, pero especialmente en los esfuerzos
gubernamentales para difundir e implicar programas de manejo como el
dar incentivos para la reforestación con especies nativas.
Como
todavía los ciclos de reproducción, requisitos
ecológicos y condiciones para una germinación óptima
de la mayoría de especies nativas estan desconocidas, mas
estudios aplicados en esta dirección estan inevitables para
dar una base sólida a cualquiera programa de reforestación.
Uso de productos
no-maderables
Por tener éxito,
planes de manejo sustentable y conservación tienen que incluir
las comunidades locales, y sus conocimientos tradicionales.
Decisiones de manejo necesitan que incluir la importancia alta de los
recursos naturales en forma de materiales de construcción,
comida, forraje y medicina, por poblaciones rurales y también
urbanas. Por esta razón, la biodiversidad andina tiene un
papel sumamente importante por la vida de muchas comunidades, y
representa una parte integral de su diversidad cultural.
El uso tradicional
de recursos forestales y alto andinas tiene raíces muy
profundos. Productos naturales, y entre ellos especialmente plantas
medicinales, tienen un papel inmensamente importante en la vida
diaria de una parte grande de la población global. Plantas
medicinales no solo representan con frecuencia la única
alternativa medicinal viable - por la baja disponibilidad y los
costos altos de la medicina del oeste, sino su colecta y venta, en
muchos casos, representan una de las oportunidades mas importantes
por poblaciones rurales, de recibir recursos monetarios. El
conocimiento sobre el uso de un recurso se pasa de frecuencia solo de
una generación a otra. Especialmente el uso de plantas
medicinales esta altamente dinámico, evolviendo continuamente,
con el descubrimiento de nuevos conocimientos y su enlace a practicas
tradicionales.
En
el Sur de Ecuador se registraron hasta ahora 142 plantas medicinales
diferentes de 64 familias (Fig. 4, Béjar et al. 2001). La
familia mas importante son las Asteraceas, seguido por Lamiaceas. De
todas las plantas, 39 familias estaban representados por una sola
especie. Este muestra claramente, que la medicina tradicional hace
uso de una gran parte de la biodiversidad, y que no se concentra solo
en un par de familias de plantas. No obstante, solo 70% de las
especies usadas son indígenas de la zona, mientras una tercera
parte representa especies introducidas, por mayoría de Europa
(Fig. 5). Este indica, que la tradición curanderistica esta
siempre en flujo - incorporando plantas nuevas si tienen
utilidad, y posiblemente descartando otras. En Ecuador ya se puede
observar una muy profunda perdida de conocimiento. Este se muestra en
la observación, que en el Norte de Perú se usa todavía
casi 700 plantas medicinales diferentes - casi 5 veces mas
especies que en Ecuador.
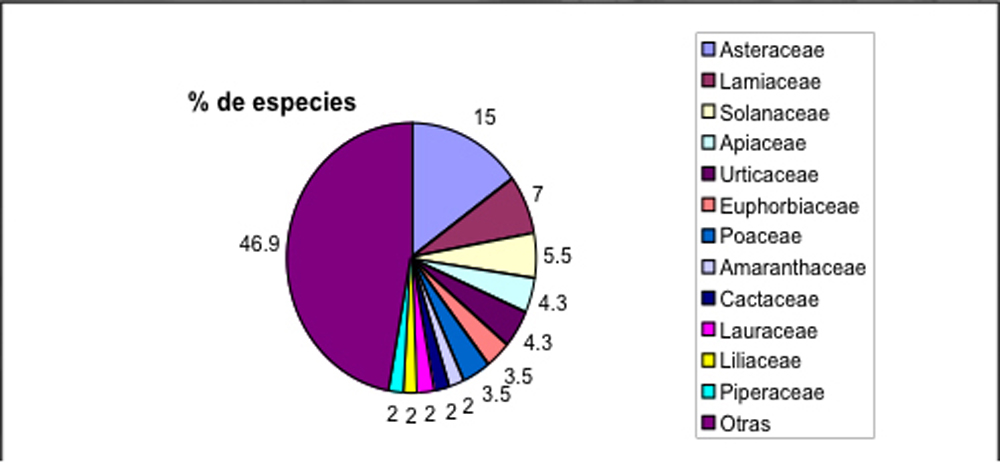
Fig. 4: Familias de plantas medicinales usadas en el
Sur de Ecuador.
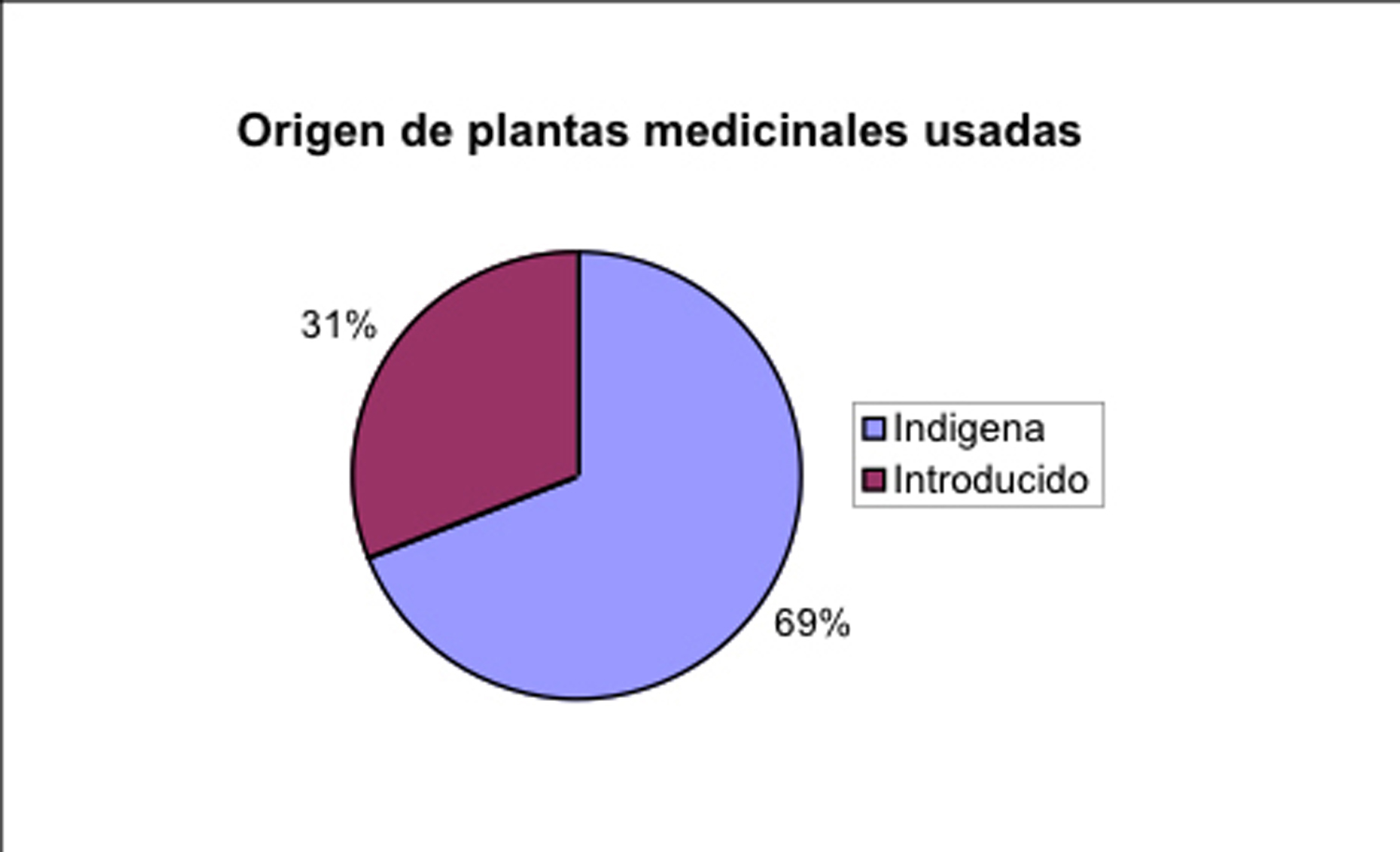
Fig. 5: Origen de plantas medicinales usadas en el Sur
de Ecuador.
Por la mayoría
de los tratamientos se usa las partes verdes de las plantas (Fig. 6),
y el material esta cocinado, que representa preparado en forma de una
tizana (Fig. 7).
El enfoque mayor de
las curaciones esta en el tratamiento de enfermedades psicosomáticos,
clasificado en manera popular como "mágicos" -
susto, mal aire, espanto, mal ojo, envidia. Mas que 35% de las
plantas están dedicadas a este uso. Enfermedades reumáticas,
problemas del sistema respiratorio, y tracto urinario representan las
categorías de enfermedad "normales" mas
importantes (Fig. 8).
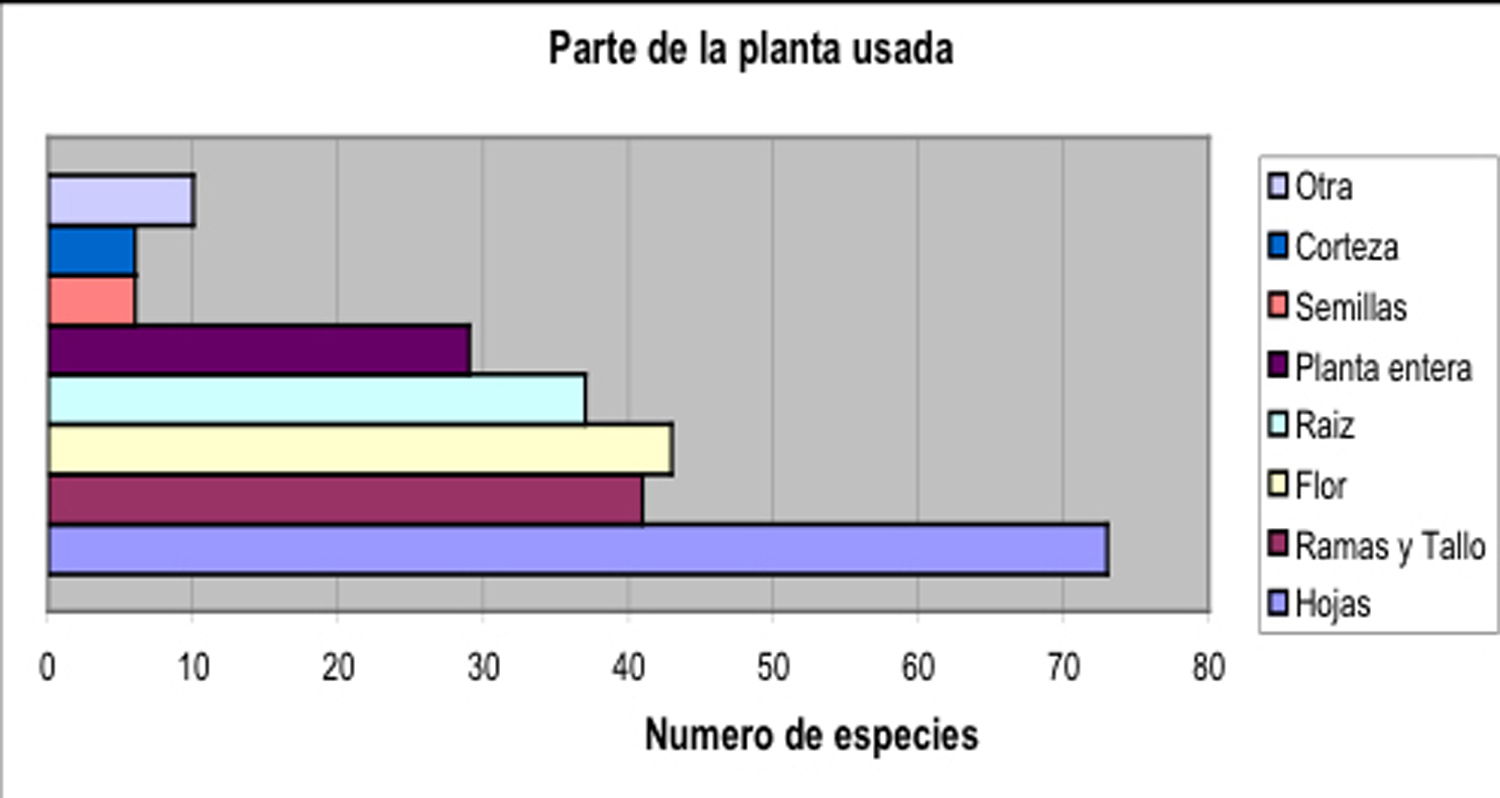
Fig. 6: Partes de plantas medicinales usadas en el Sur
de Ecuador.
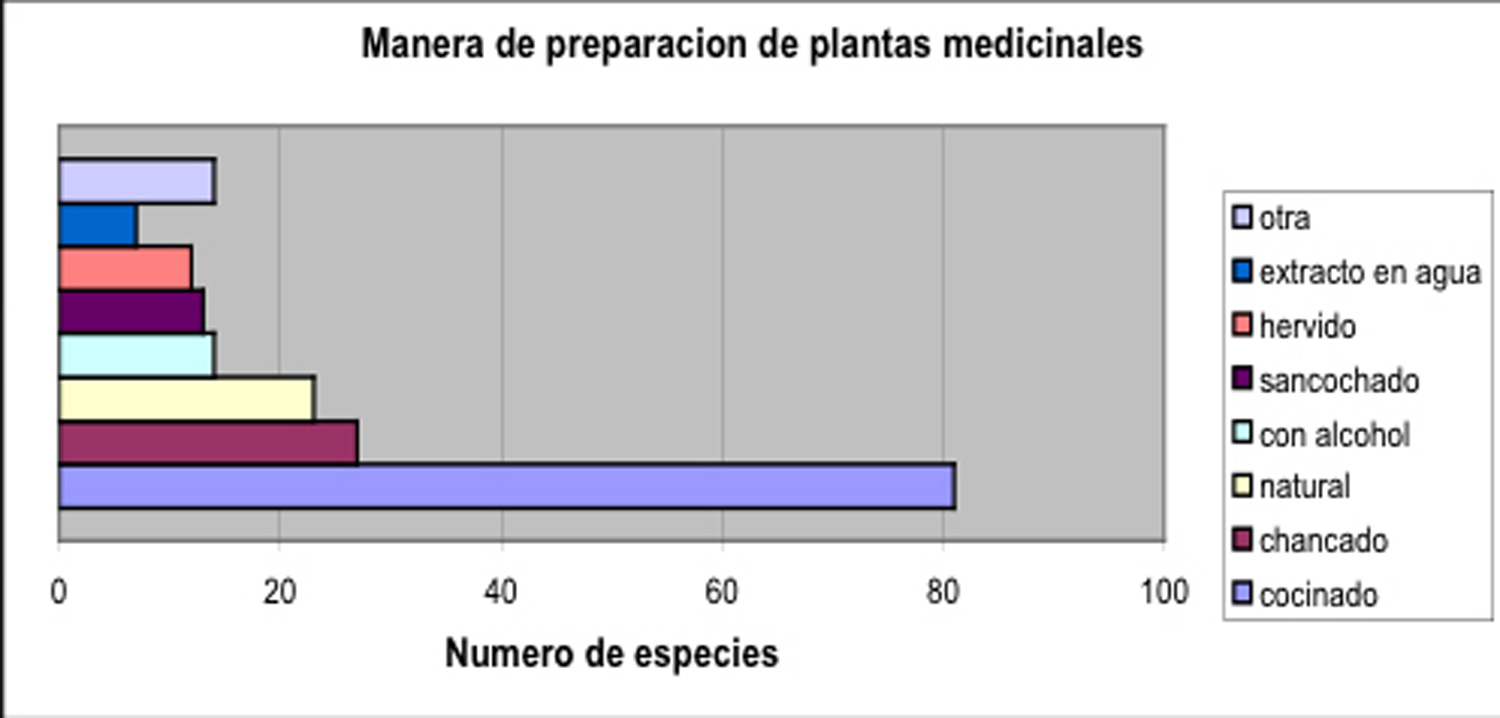
Fig. 7: Preparación de plantas medicinales
usadas en el Sur de Ecuador.
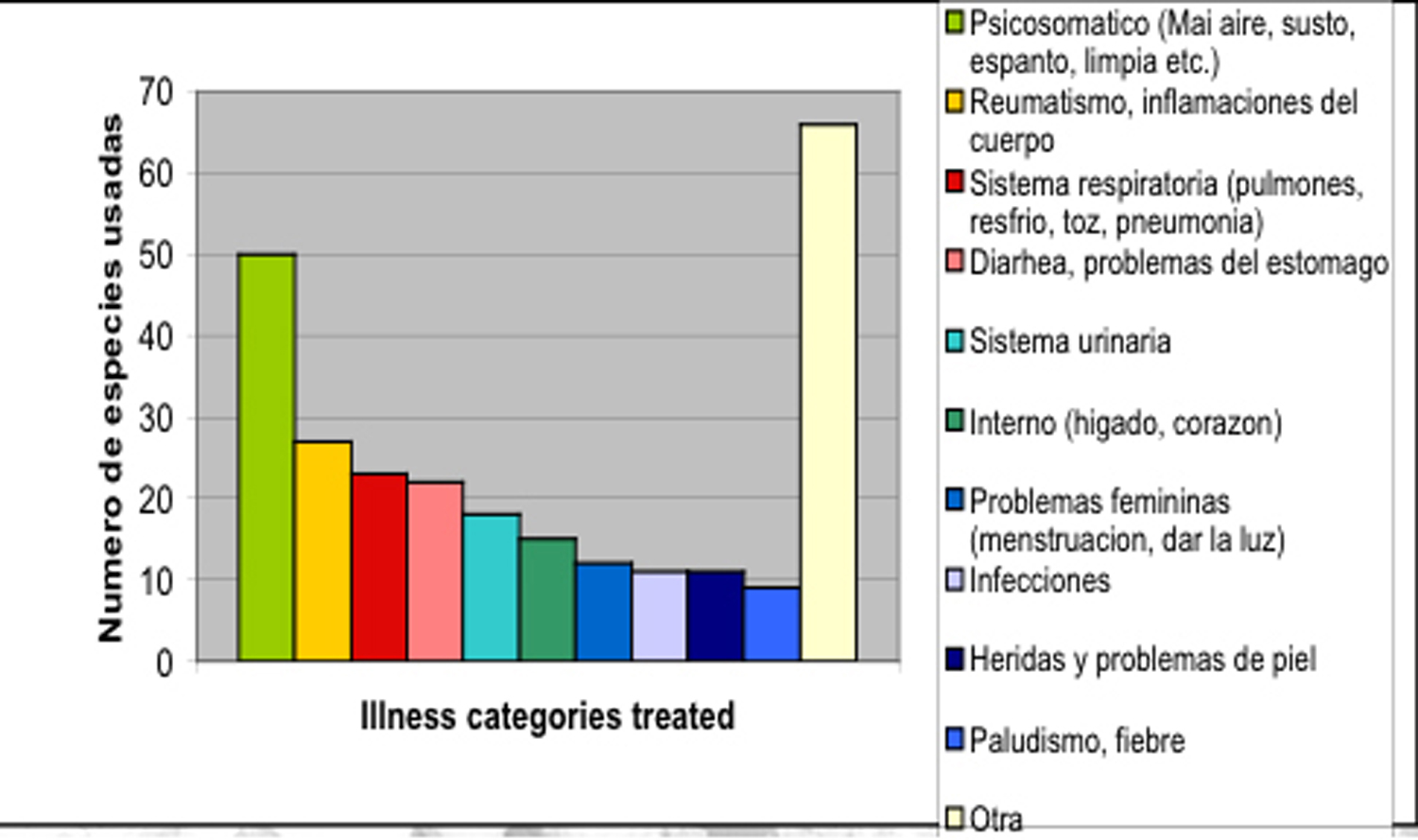
Fig. 8: Clases de enfermedades curadas con palntas
medicinales en el Sur de Ecuador.
Los
posibles beneficios económicos del uso de recursos medicinales
de la selva y región altoandina, en vez de la cosecha de solo
productos maderables, parece muy interesante por un mercado local.
Sin embargo, estos mismos recursos se encuentran inmensamente
amenazados por tala indiscriminada, quema de bosque y minería
(Fig. 9). Estas amenazas, si siguen sin control, representan un
peligro enorme por la diversidad biológica y cultural andina.

Fig. 9: Destrucción de Biodiversidad.
Extracción de Madera, Ecuador (izquierda arriba, derecha
abajo), Quema del bosque, Ecuador (derecha arriba), Minería,
Ecuador (izquierda abajo).
Discusión
Los
bosques encontrados en El Sur de Ecuador y Norte de Perú se
agrupan en tres formaciones. Como la diversidad de bosques montanos
consiste por gran parte por hierbas, arbustos y epifitas (Gentry
1988, 1995; Gentry & Dodson 1987), mientras árboles tienen
un papel menos importante que en bosques húmedos tropicales,
una metodología enfocando no solo en especies leñosas
grandes que permite un entendimiento mejor de la diversidad de estos
bosques. Los bosques montanos estudiados muestran diferencias
profundas en comparación a otros áreas. La abundancia
muy alta de Alzathea verticillata (Alzatheaceae) - nunca
antes registrado en Ecuador (Foster comm. pers.) esta única,
aunque en general la composición florística se puede
comparar a otros áreas (Frei 1958, Grubb et al. 1963, 1966;
Madsen 1989, 1991; Madsen & Øllgaard 1994). En lo
contrario el Bosque montano alto, dominado por Purdiaea nutans,
es un caso muy especial de una formación de bosque muy
aislada, con solo unosáreas en el Norte de Perú
comparecidos. En Cajanuma, aún muy cerca, la composición
de los bosques esta completamente diferente (Madsen 1989, 1991;
Madsen & Øllgaard 1994). En manera florística los
bosques de la ECSF no están representativos para un bosque
andino del Sur de Ecuador, y menos para los bosques andinos en
general.
Atentos
de conservar y manejar la diversidad biológica tienen que
integrar los puntos de vista, las necesidades y el conocimiento
tradicional de las poblaciones locales y indígenas. El
intercambio entre científicos, ONGs y comunidades locales esta
inevitable en este proceso, y cada parte de esta red necesita
contribuir al éxito. En el nivel científico esta de
mayor importancia de incluir estudios básicos sobre ecología
o etnobotanica en un ambiente amplió, holistico,
interdisciplinario, para elaborar una base estable por la
implementación de los resultados. Estos deben ser disponibles
en el idioma local - en una manera entendible y practica. A
largo plazo, solo manejo y conservación basado en trabajos
holisticos vaya tener éxito.
Agradecimientos
Agradezco mucho el
apoyo continuo de la Deutsche Forschungsgemeinschaft (Fundación
Alemana para la Investigación - DFG) en el margen del
proyecto "Funcionalidad en un bosque montano tropical:
Diversidad, procesos dinámicos y potencial para el uso"
(BE 473/28-1,2,3; Bu 886/1-1,2,4, FOR 402-1/TP7) y el programa MIRT
del Instituto Nacional de Salud (NIH). Quiero agradecer en manera
especial el apoyo de mis contrapartes Peruanos y Ecuatorianos, en
particular de los equipos de TRUX y HUT.
Referencias
Bgh,
A. 1992. Composition and distribution of the vascular epiphyte flora
of an Ecuadorian montane rainforest. - Selbyana 13: 25-34.
Braun-Blanquet,
J. 1964. Pflanzensoziologie (3rd edition). Wien, New York.
Bussmann,
R.W. 1999. Forest vegetation units along altitudinal gradients at
Estación Científica San Francisco, Ecuador.
Abstract 12. Jahrestagung der Deutschen Gesellschaft für
Tropenökologie. Ulm/Donau.
Bussmann,
R.W. 2001. The montane forests of Reserva Biológica San
Francisco (Zamora-Chinchipe, Ecuador) vegetation zonation and natural
regeneration. - Die ERDE 132: 11-24.
Bussmann,
R.W. & S. Lange 1998. Expedición Rapido Asesso
Cordillera Sabanilla - Inventario floristico. INEFAN, Loja/Quito.
Churchill,
S.P., H. Balslev, E. Forero, & J.L. Luteyn, (eds.) 1995.
Biodiversity and Conservation of Neotropical Montane Forests.
NYBG: New York. 703pp.
Cleef,
A.M. 1981. The Vegetation of the Páramos of the Colombian
Cordillera Oriental. - Dissertationes Botanicae 61. Cramer:
Vaduz.
Ek,
R.C. 1997. Botanical diversity in the tropical rain forest of
Guayana. - Tropenbos-Guayana series 4: 1-237.
Emck,
P. en prep. Climatic conditions in the Cordillera de Numbala
(Podocarpus National Park), Southern Ecuador. PhD tesis,
University of Erlangen.
Emperaire,
L. & C. Friedberg, 1990. Relevées floristiques des
regions Piura (Perou) et de Loja (Ecuateur). ORSTROM, Paris.
Espinosa,
B. 1948a. Estudios botanicos en el sur del Ecuador. I.
Loja-Catamayo-Malacatos-Vilcabamba. Loja, Universidad Nacional.
Espinosa,
B. 1948b. Estudios botanicos en el sur del Ecuador. II. Herbarium
Universitatis Loxoensis (Primer Inventario). Loja, Universidad
Nacional.
Espinosa,
G.D.A. (ed.) 1989/1992. Parque Nacional Podocarpus. Boletin
informativo sobre biologia, conservacion y vida silvestre 1-3.
Universidad Nacional: Loja.
Frei,
E. 1958. Eine Studie ¸ber den Zusammenhang zwischen Bodentyp,
Klima und Vegetation in Ecuador. - Plant and Soil 9: 215-236
Gálvez
M., J.R., Ordoñez
G., O.R. & Bussmann, R.W. 2003.
Estructura del bosque montano perturbado y no-perturbado en el Sur
de Ecuador. Lyonia 3(1): 83-98.
Garwood,
N.C. 1985. Earthquake-caused landslides in Panama: Recovery of
vegetation. - Res. Rep. Natl. Geogr. Soc. 21: 181-184.
Gentry,
A.H. & C.H. Dodson 1987. Contribution of nontrees to species
richness of a tropical rain forest. - Biotropica 19: 149-156.
Gentry,
A.H. 1988: Changes in plant community diversity and floristic
composition on environmental and geographical gradients. - Ann.
Missouri Bot. Gard. 75: 1-34.
Gentry,
A.H. 1995. Patterns of diversity and floristic composition in
neotropical montane forests. - In: Churchill, S.P., H. Balslev, E.
Forero, & J.L. Luteyn (eds.): Biodiversity and conservation of
Neotropical montane forests, pp. 103-126. - NYBG, New York.
Grubb,
P.J. &. T.C. Whitmore 1966. A comparison of montane and lowland
rain forest in Ecuador - II. The climate and its effects on the
distribution and physiognomy of the forests. - J. Ecol. 54:
303-333.
Grubb,
P.J., J.R. Lloyd, T.D. Pennington, & T.C. Whitmore 1963. A
comparison of montane and lowland rain forest in Ecuador - The forest
structure, physiognomy, and floristics. - J. Ecol. 51:
567-601.
Guariguata,
M.R. 1990. Landslide disturbance and forest regeneration in the upper
Luquillo mountains of Puerto Rico. - J. Ecol. 78: 814-832.
Hall,
M. 1977. El volcanismo en el Ecuador. Abya Yala: Quito.
Hamilton,
L.S., J.O. Juvik y F.N. Scatena (eds.). 1994. Tropical Montane Cloud
Forests. Ecological Studies 110: New York.
Hammen,
T. van der & A.G. Dos Santos, (eds.) 1995. Studies on tropical
Andean ecosystems Vol. 4. Cramer: Vaduz. 603pp.
Hammen,
T. van der & P.M. Ruiz, (eds.) 1984. Studies on tropical
Andean ecosystems Vol. 2. Cramer: Vaduz. 603pp.
Hammen,
T. van der, A. Perez Precario y E.P. Pinto (eds.) 1983. Studies on
tropical Andean ecosystems Vol. 1. Vaduz.
Hammen,
T. van der, D. Mueller-Dombois y M.A. Little 1989. Manual of
Methods for Mountain Transect Studies. Paris.
Hammen,
T. van der, S. Diaz-Piedrahita y V.J. Alvarez (eds.) 1989. Studies
on tropical Andean ecosystems Vol. 3. Vaduz.
Herzog,
T. 1923. Die Pflanzenwelt der bolivianischen Anden und ihres
oestlichen Vorlands. Leipzig.
Jrgensen,
P.M. & C. Ulloa Ulloa 1994. Seed plants of the High Andes of
Ecuador - a checklist. - AAU Reports 34: 1-443.
Jrgensen,
P.M. & S. León-Yanez, (eds.) 1999. Catalogue of the
vascular plants of Ecuador. - Monographs in Systematic Botany from
the Missouri Botanical Garden 75.
Jrgensen,
P.M. 1991. Species composition and present extension of Andean
forest above 2,400 m altitude in Ecuador. - Ph.D. thesis,
Botanical Institute of Aarhus University, unpublished.
Kessler,
M. 1999. Plant species richness and endemism during natural landslide
succession in a perhumid montane forest in the Bolivian Andes. -
Ecotropica 5(2). 123-136.
Leischner,
B. & R.W. Bussmann. 2003. Mercado
y uso de madera en el Sur de Ecuador. Lyonia 5(1):
51-60.
Madsen,
J.E. & B. Øllgaard 1994. Floristic composition, structure
and dynamics of an upper montane rain forest in Southern Ecuador. -
Nord. J. Bot. 14(4): 403-423.
Madsen,
J.E. 1989. Aspectos generales de la flora y vegetación del
Parque Nacional Podocarpus. - Boletin informativo sobre biología,
conservación y vida silvestre 1: 59-74.
Madsen,
J.E. 1991. Floristic composition, structure and dynamics of an
upper montane rain forest in Southern Ecuador. - Ph.D. thesis,
Botanical Institute of Aarhus University, unpublished.
Meier,
W. 1998. Flora und Vegetation des Avila-Nationalparks Venezuela/
Kuestenkordillere) unter besonderer Ber¸cksichtigung der
Nebelwaldstufe. - Dissertationes Botanicae 296. Cramer:
Berlin.
Mueller-Dombois,
D. & H. Ellenberg 1974. Aims and methods of vegetation
ecology. Wiley: New York.
Øllgaard,
B. & J.E. Madsen 1993. Inventario preliminar de las especies
vegetales en el Parque Nacionál Podocarpus. - Revista de
Difusión Técnica y Cientifica de la Facultad de
Ciencias Agrícolas, Universidad Nacional de Loja 22/23
(1/2): 66-87.
Schrumpf,
M., G. Guggenberger, C. Valarezo & W. Zech 2001. Tropical rain
forest soils. Die ERDE 132: 43-59.
Sierra,
R. (ed.) 1999. Propuesta preliminar de un sistema de clasificación
de vegetación para el Ecuador Continental. GEF: Quito,
194pp.
Simonett,
D.S. 1967. Landslide distribution and earthquakes in the Bewani and
Torricelli mountains, New Guinea. - In: Jennings, J.N. & J.A.
Mabbutt (eds.): Landform studies from Australia and New Guinea,
pp. 64-84. - Canberra.
Stern,
M.J. 1995. Vegetation recovery on earthquake-triggered landslide
sites in the Ecuadorian Andes. In: Churchill, S.P., H. Balslev, E.
Forero & J.L. Luteyn (eds.): Biodiversity and conservation of
neotropical montane forests., 207-220. NYBG: New York.
Ulloa,
C. & P.M. Jrgenson 1993. Arboles y arbustos de los Andes
del Ecuador. - AAU Reports 30: 1-263.
15
. Vegetation recovery on earthquake-triggered landslide
sites in the Ecuadorian Andes. In: Churchill, S.P., H. Balslev, E.
Forero & J.L. Luteyn (eds.): Biodiversity and conservation of
neotropical montane forests., 207-220. NYBG: New York.
Ulloa,
C. & P.M. Jrgenson 1993. Arboles y arbustos de los Andes
del Ecuador. - AAU Reports 30: 1-263.
15