Introducción

El
género Lachemilla se distribuye en las zonas montañosas
occidentales del neotrópico, desde México hasta el
norte de Chile y Argentina, entre los 2200 y 5000 m., con una sola
especies en República Dominicana. Se han descrito cerca de 80
especies de Lachemilla, de las cuales 36 (45%) se encuentran
en la región norte de los Andes sudamericanos, desde Venezuela
hasta Ecuador.
Lachemilla
es un género ecológicamente importante ya que muchas
veces forma grandes asociaciones vegetales que actúan como
reservorios naturales de agua en sitios muy elevados como son los
páramos y superpáramos; parte del agua utilizada por
las poblaciones alto-andinas proviene de los páramos.
Aunque
Lachemilla es un género común y diverso en las
montañas neotropicales y en especial en la región
alto-andina del norte de Sudamérica, existen relativamente
pocos estudios taxonómicos sobre este género y casi
nada se conoce sobre su ecología. Este estudio es parte del
tratamiento taxonómico que se está realizando
actualmente sobre Lachemilla.
Previos estudios taxonómicos sobre
Lachemilla
La
primera y única revisión tentativa de Lachemilla
la realizó L. M.. Perry (1929) en la que reconocio 41
especies. Posteriormente, Rothmaler (1935 1937) publicó las
Lachemilla de Colombia, y además varias nuevas especies
y combinaciones, y una clasificación preliminar del género
en la que reconoce 72 especies. En 1977, E. G. B. Kieft, del herbario
de Utrecht empezó la revisión de Lachemilla pero
nunca la concluyó, aunque identificó bastante material.
Recientemente el género ha sido estudiado en Venezuela
(Gaviria 1996) y en Ecuador (Romoleroux 1996).
Delimitación de Lachemilla y
afinidades taxonómicas
Dentro
de la familia Rosaceae, Lachemilla se ubica en la subfamilia
Rosoideae y tradicionalmente en la tribu Sanguisorbeae o Poterieae
(Hutchinson 1964), junto con otros géneros como Acaena
o Polylepis que carecen de pétalos y que probablemente
presentan una tendencia a polinización por el viento. Sin
embargo, Schultze-Menz (1964), saca a Lachemilla, Aphanes y
Alchemilla de Sanguisorbeae y los ubica en la tribu
Potentilleae, estudios filogenéticos y moleculares recientes
apoyan este cambio (Ericksson et al. 2003).
Lachemilla
fue creada como una sección del género Alchemilla
(Focke 1888), (Perry 1929); sin embargo, Rydberg (1908) y Rothmaler
(1937) lo trataron como un género distinto. Lachemilla
y Alchemilla comparten algunas características
morfológicas, pero difieren en varios aspectos, como son: la
distribución geográfica, el número de estambres
y la posición de los estambres y anteras.
Clasificación infragenérica
El
género Lachemilla es morfológicamente muy
diverso y ha sido subdividido en grupos infragenéricos. Perry
(1929) reconocio series y seis subseries; mientras Rothmaler (1937)
publicó seis secciones y seis subsecciones. Las dos
clasificaciones coinciden en que el grupo ancestral es L.
polylepis, pero según Perry la especie más derivada
es L. diplophylla y según Rothmaler es el grupo de L.
aphanoides.
En
el presente trabajo se proporciona una descripción de
Lachemilla, una clave de las especies para Ecuador, Colombia y
Venezuela, y una lista de las especies en estos tres países
con su distribución y hábitats.
Descripción de Lachemilla y
caracteres taxonómicos utilizados para la identificación
El
género Lachemilla comprende hierbas perennes o pocas
veces pequeños arbustos. Las hojas son simples, lobuladas,
tripartidas o pinnaticompuestas, con estípulas. Las flores se
disponen en cimas glomeruladas o laxas o son solitarias. Cada flor
presenta un hipantio que es una prolongación del receptáculo,
en el que están insertos el cáliz, epicáliz y
los estambres, carece de pétalos; el cáliz tiene 4
sépalos, el epicáliz es un verticilo exterior al cáliz,
generalmente con 4 episépalos (a veces 2 o ausentes) alternos
a los sépalos; los estambres son 2 o raramente 3 o 4 y están
en el borde interno del disco del hipantio, las anteras son
extrorsas; el gineceo puede tener uno a 10 carpelos encerrados en el
hipantio, cada carpelo presenta un óvulo basal. El fruto es
seco e indehiscente con uno a 10 aquenios.
Los caracteres
taxonómicos más utilizados para la identificación
de las especies son:
Hábito:
generalmente son herbáceas pequenãs, decumbentes,
reptantes, estoloníferas o ascendentes; solo una especie es
completamente arbustiva y leñosa: Lachemilla polylepis,
que es considerada como la más ancestral dentro del género.
Pubescencia: todas
las especies presentan algun tipo de pubescencia, es un carácter
útil para la identificación de especies.
Estípulas:
la forma y textura de las estípulas es variable, están
adnadas parcialmente a la base del pecíolo y libres al ápice,
o pueden estar adnadas y connadas hacia la base, con lóbulos
libres hacia el ápice formando verticilos. La textura es
normalmente herbácea, al menos hacia el ápice, solo
tres especies tienen estípulas completamente papiráceas:
L. polylepis, L. tanacetifolia y L. rupestris.
Hojas:
La forma de las hojas es extremadamente variable, en la misma planta,
las hojas basales y distales son usualmente diferentes. En varias
especies las hojas presentan un pecíolo definido y una lámina
que puede ser: simple lobulada e.g. L. orbiculata ([[Figura
1]]), 3-5 partida e.g. L. hirta ([[Figura 2]]) o pinnada e.g.
L. pinnata ([[Figura 3]]); en otro grupo de especies, los
pecíolos están reducidos o ausentes y las láminas
de las hojas se reducen a simples lóbulos iguales a los
lóbulos de las estípulas y forman un verticilo
alrededor de las ramas, e.g. L. nivalis ([[Figura 4]]). El
caso más extremo de modificación de las hojas es el de
L. diplophylla ([[Figura 5]]) que presenta una lámina
ligeramente bilobada y dos apéndices foliares que nacen
ventralmente a lo largo del nervio principal, dando la apariencia de
doble hoja.
Inflorescencia:
las flores están agrupadas en cimas, laxas o glomeruladas;
pocas especies presentan flores solitarias, e.g. L. uniflora
o L. diplophylla ([[Figura 5]]), éste se considera
un estado más evolucionado dentro del género.
Hipantio:
la forma y la pubescencia del hipantio son útiles para la
identificación, el hipantio puede ser externamente pubescente
o grabrescente, e internamente glabro, solo un grupo de especies
presenta la parte interna del hipantio pubescente, e.g. L. hirta
([[Figura 2]]).
Cáliz
y epicáliz: el tamaño del cáliz en relación
al epicáliz es útil para diferenciar algunas especies;
el epicáliz está presente en la mayoria de especies
pero en algunas es muy reducido o incluso completamente ausente, e.g.
L. diplophylla ([[Figura 5]]).
Carpelos y frutos:
El número de carpelos es, a veces, útil para reconocer
algunas especies, al igual que el número y la forma de los
aquenios.
Clave
de las especies de Lachemilla en el norte de los Andes
Sudamericanos (Venezuela, Colombia y Ecuador)
1a. Arbustos, tallo
y ramas completamente leñosos, estípulas papiráceas
formando vainas persistentes alrededor del tallo. 1. L. polylepis
1b. Hierbas o raro
subarbustos, tallo herbáceo o parcialmente lignificado, sin
estípulas papiráceas formando vainas persistentes
alrededor del tallo
2a. Hojas lobuladas,
partidas o pinnadas, sin apéndices.
3a. Estípulas
distales enteras, completamente papiráceas.
4a. Hojas 3-partidas
2. L. rupestris
4b. Hojas
pinnadas 3. L. tanacetifolia
3b. Estípulas
distales lobuladas o partidas, herbáceas por lo menos hacia el
ápice.
5a. Inflorescencias
cimas laxas; superficie interna del hipantio pubescente.
6a. Hojas 3-partidas
con segmentos laterales bífidos; largo de los lóbulos
estipulares 2/5 a 1/2 de la longitud total de la vaina estipular.
7a. Ramas hirsutas;
episépalos más largos que los sépalos. Hojuelas
espatuladas a elongado-obovadas 4. L. hirta
7b. Ramas
subsericeas, pilosas o glabrescentes; episépalos más
cortos o iguales que los sépalos. Hojuelas mayormente
obovadas 5. L. andina
6b. Hojas 3-partidas
con segmentos laterales enteros; largo de los lóbulos
estipulares 3/5 a 3/4 de la longitud total de la vaina estipular.
8a. Ramas
esparcidamente villosas hasta glabrescentes; lóbulos
estipulares a menudo reflexos, lóbulos foliares profundamente
dentados 6. L. vulcanica
8b. Ramas sericeas;
lóbulos estipulares ascendentes, lóbulos foliares
aserrado-dentados
7. L. jamesonii
5b. Inflorescencias
cimas densas, glomeruladas o flores solitarias; superficie interna
del hipantio glabra.
9a. Hojas distales
reducidas, iguales o casi iguales a los lóbulos estipulares,
adnadas y connadas a las estípulas formando muchas vainas
perfoliadas a lo largo del tallo.
10a. Hojas distales
iguales a los lóbulos estipulares.
11a. Lóbulos
de las vainas a lo largo del tallo 4-6
12a. Lóbulos
de las vainas distales glabros (Venezuela) o esparcidamente villosos
(Colombia y Ecuador) y reticulados en el enves 8. L. equisetiformis
12b. Lóbulos
de las vainas distales hispidulos y no reticulados en el enves 9.
L. ericoides
11b. Lóbulos
de las vainas a lo largo del tallo 8-12
13a. Lóbulos
de las vainas ascendentes. Ramas pilosas, villosas o sericeas; hojas
basales caducas
14a. Largo de los
lóbulos de la vaina más de 4/5 de la longitud total de
la vaina; lóbulos de la vaina 7 a 8 veces más largos
que anchos 10. L. angustata
14b. Largo de los
lóbulos de la vaina más de 2/5 de la longitud total de
la vaina; lóbulos de la vaina 2 a 4 veces más largos
que anchos
15a. Diámetro
de los tallos más de 5 mm, villosos, pilosos o glabrescentes.
Sépalos y episépalos conniventes 11. L. nivalis
15b. Diámetro
de los tallos menos de 4 mm, sericeos. Sépalos y episépalos
rectos
12. L. ramosissima
13b.
Lóbulos de las vainas ligeramente ascendentes a horizontales o
reflexos. Ramas híspidas, hirsutas o villosas; hojas basales
usualmente presentes (excepto en L. hispidula).
16a. Lóbulos
de las vainas 9-13; ramas híspidas o hirsutas; hipantio glabro
hacia la base
17a. Ramas hirsutas,
lóbulos de las vainas reflexos, lanceolados 13. L. galioides
17b. Ramas híspidas,
lóbulos de las vainas ligeramene ascendentes a horizontales,
deltoide-subulados
18a. Plantas
ascendentes, usualmente más de 12 cm de largo, hojas basales
frecuentemente ausentes, brácteas florales ascendentes 14.L.
hispidula
18b. Plantas
decumbentes, menos de 10 cm de largo, hojas basales presentes,
brácteas florales horizontales 15. Lachemilla sp. Nov.
16b. Lóbulos
de la vaina 7-8 (10); ramas villosas; hipantio completamente sericeo
o villoso 16. L. verticillata
10b. Hojas distales
ligeramente más largas que los lóbulos estipulares,
3-partidas o 3-lobuladas.
19a.
Hojas basales a menudo caducas; episépalos más largos o
iguales a los sépalos; carpelos ≥ 6.
20a. Planta sericea
o villosa-sericea, lóbulos foliares y estipulares cartáceos
(Colombia, Ecuador y Perú) 17. L. holosericea
20b.
Planta estrigosa a glabrescente, lóbulos foliares y
estipulares coriaceos (Venezuela) 18. L. trevirani
19b. Hojas basales
persistentes; episépalos más cortos que los sépalos;
carpelos ≤ 6.
21a.
Hojas basales 5-partidas; hojas distales 3-partidas, segmentos
inciso-dentados, segmento central 3-lobulado. Tallos ascendentes a
erectos 19. L. sprucei
21b. Hojas basales
3-partidas; hojas distales 3-partidas, segmentos laterals usualmente
enteros, segmento central 3-lobulado. Tallos procumbentes,
decumbentes a ± ascendentes.
22a. Plantas
villosas o hirsuta-villosas, largo de los lóbulos de las
estípulas menos de 2/4 de la longitud total de la vaina. 20.
L. holmgrenii
22b. Plantas
sericeas o glabrecentes, largo de los lóbulos de las estípulas
más de 2/4 de la longitud total de la vaina.
23a. Plantas
glabrescentes, con glándulas en sépalos, episépalos
y hacia la base de los tricomas 21. L. rivulorum
23b. Plantas
sericeas o esparcidamente sericeas, sin glándulas.
24a. Tallos ±
ascendentes, nudos de las ramas sin raíces 22. L. killipii
24b. Tallos
reptantes a procumbentes, nudos de las ramas usualmente con raíces
23. L. purdiei
9a. Hojas distales
desarrolladas (si son reducidas entonces las hojas basales
orbiculadas), lóbulos foliares más largos que los
lóbulos estipulares.
25a. Hojas basales
orbicular-reniformes, ligeramente (7—) 9—11 lobuladas.
26a. Brácteas
florales connadas, envuelven parcialmente a las flores; margen de las
hojas crenado-aserrados; hojas distales usualmente ausentes; carpelos
2-4. 24. L. orbiculata
26b. Brácteas
florales libres, no envuelven a las flores; margen de las hojas
pectinados; hojas distales presentes; carpelos 4-6. 25. L.
pectinata
25b. Hojas basales
5-lobuladas, 3-partidas o pinnati-compuestas.
27a. Hojas basales
profundamente 5-lobuladas o 3-partidas.
28a. Hojas basales
5-lobuladas, persistentes
29a. Episépalos
y sépalos abaxialmente glabros, ascendentes o reflexos; flores
menos de 2 mm de largo; margenes de las hojas aserrados o ligeramente
subincisos 26. L. aequatoriensis
29b. Episépalos
y sépalos abaxialmente pubescentes, rectos; flores más
de 2 mm de largo; margenes de las hojas incisos 27. L. fulvescens
28b. Hojas basales
3-partidas o caducas
30a. Hojas basales y
caulinares 3-partidas, persistentes; plantas procumbentes o
estoloníferas
31a. Flores en cimas
glomeruladas, generalmente más de 5 flores
32a. Envés de
las hojas esparcidamente piloso o sericeo-villoso, no glauco; margen
glabro; hipantio externamente pubescente; carpelos 4-5.
28. L. perryana
32b. Envés de
las hojas esparcidamente hirsuto-villoso, glauco; margen ciliado;
hipantio externamente glabrescente; carpelos 6-7 29. L. mutellina
31b. Flores
solitarias o en cimas axilares, paucifloras, menos de 5 flores. 30.
L. uniflora
30b. Hojas basales
caducas, caulinares 3-partidas; plantas decumbentes.
33a. Hipantio glabro
o ± piloso; segmentos foliares laterales no bifidos, margen
inciso. 31. L. aphanoides
33b. Hipantio
sericeo o adpreso-hirsuto; segmentos foliares laterales bifidos,
margen dentado-aserrado 32. L. moritziana
27b. Hojas basales
pinnadas
34a. Episépalos
presentes
35a. Hojas basales
linear-ovadas en contorno, 3 o más veces más largas que
anchas, con 6-10 pares de hojuelas 33. L. pinnata
35b. Hojas basales
deltoide-ovadas en contorno, menos de 2 veces más largas que
anchas, con 2-3 pares de hojuelas
36a. Pedicelos menos
de 2 mm de largo; hojas bipinnadas 34. L. erodiifolia
36b. Pedicelos más
de 2 mm de largo; hojas pinnadas 35. L. paludicola
34b. Episépalos
ausentes 35. L. paludicola
2b. Hojas
ligeramente bilobuladas, con 2 apéndices foliares a lo largo
del nervio principal del haz de la lámina 36. L.
diplophylla

Fig. 1. L.
orbiculata
Fig. 1. L.
orbiculata.
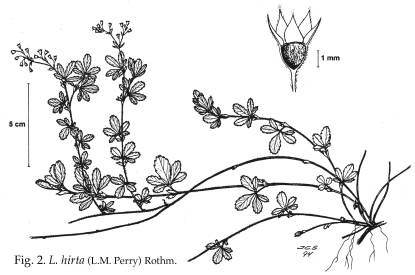
Fig. 2. L. hirta
Fig. 2. L. hirta.
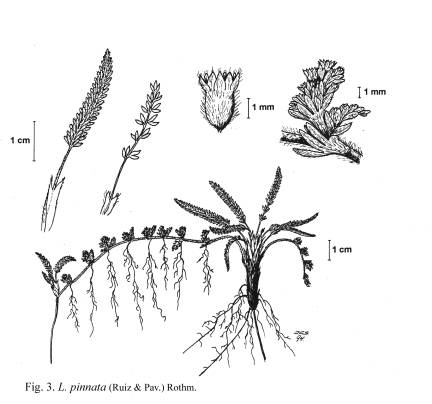
Fig. 3. L. pinnata
Fig. 3. L.
pinnata.
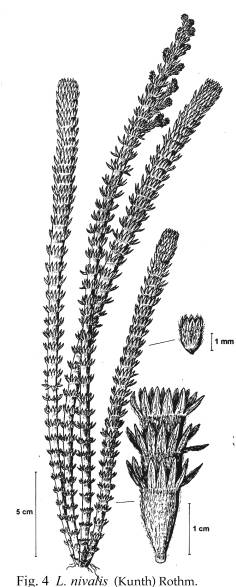
Fig. 4. L. nivalis
Fig. 4. L.
nivalis.
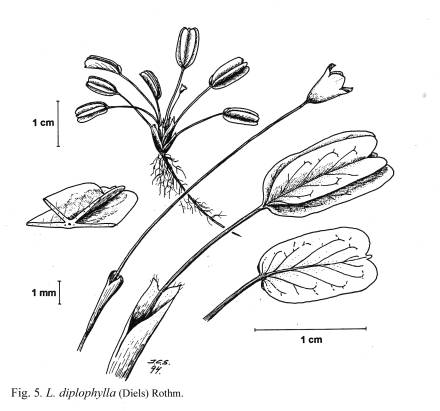
Fig. 5. L.
diplophylla
Fig. 5. L.
diplophylla.
NOTA: Las figuras
fueron tomadas del tratamiento de la familia Rosaceae para la Flora
of Ecuador, Vol. 56. Nos hemos puesto de acuerdo con los editores de
la Flora para usar estos dibujos en este artículo.
Distribución, diversidad y hábitats
La
distribución total de Lachemilla, desde el sur de
California hasta el norte de Chile y Argentina, de dividió en
tres zonas ([[Figura 6]]). Esta división se basó
principalmente en que la zona de páramo del norte de los
Andes, desde Venezuela hasta Ecuador, ha sido identificada como un
ecosistema característico (Luteyn 1999). Por lo tanto, se
trató esta parte como la zona central, desde Panamá
hasta el sur de California como la zona norte y desde Perú
hasta el norte de Chile y Argentina como zona sur. La diversidad
máxima de Lachemilla está en la zona central con
36 especies, de las cuales 22 son endémicas, el número
de especies disminuye hacia el norte y hacia el sur.
En
cuanto a los hábitats, algunas especies de Lachemilla,
crecen solo en partes muy húmedas, como es el caso de: L.
paludicola y L. diplophylla. Sin embargo, la mayoría
de especies se encuentran en diferentes tipos de páramos o en
el bosque alto-andino ([[Tabla 1]]). Al comparar los hábitats
de las especies consideradas más ancestrales y más
derivadas según los tratamientos taxonómicos anteriores
(Perry 1929; Rothmaler 1937), no se ha encontrado una relación
directa.
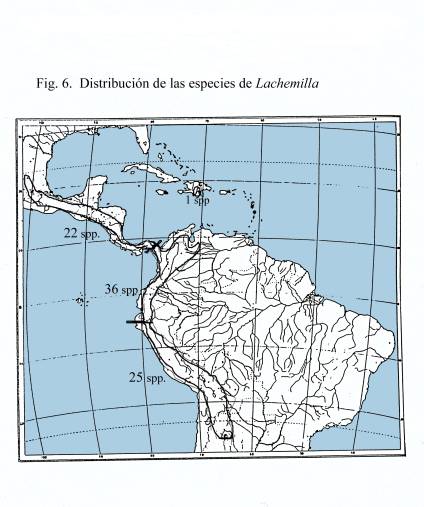
Fig. 6.
Distribución de Lachemilla
Fig.
6. Distribución de Lachemilla.
En este estudio no
se proporciona aún una clasificación infragenérica,
ya que la revisión del género no se ha concluido; sin
embargo, parte del material estudiado presenta una variación
tan gradual que las diferencias entre algunos de los grupos
infragenéricos propuestos previamente (Perry 1929, Rothmaler
1937) parecen muy artificiales.
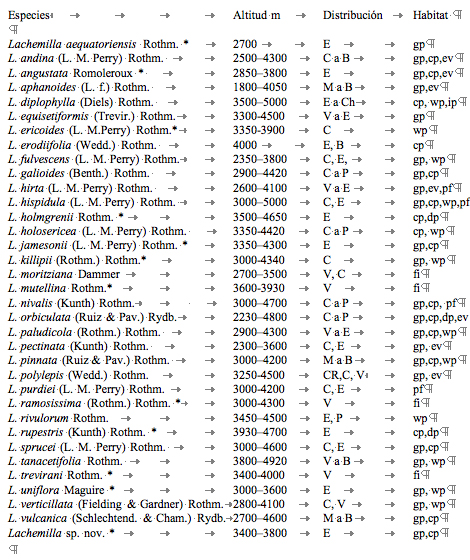
Tabla 1.
Distribución y hábitats de las especies de Lachemilla
en Venezuela Colombia y Ecuador.
Distribución:
B= Bolivia; C= Colombia; Ch= Chile; CR= Costa Rica; E= Ecuador; M=
México; P= Perú; V= Venezuela. Habitat: gp= páramo
de pajonal; cp= páramo arbustivo y de almohadilla; dp= páramo
seco; ip= páramo inundado; wp= páramo húmedo;
ev= vegetación abierta en zona de bosque y áreas
cultivadas; pf= bosque de Polylepis; fi= falta información.
*= especies endémicas.
Table
1. Distribution and habitats of Lachemilla species in
Venezuela, Colombia and Ecuador.
Distribution:
B= Bolivia; C= Colombia; Ch= Chile; CR= Costa Rica; E= Ecuador; M=
México; P= Peru; V= Venezuela. Habitat: gp=
grass paramo; cp= cushion and shrubby paramo; dp= desert paramo; ip=
flooded paramo; wp= wet paramo; ev= exposed vegetation in the forest
zone and cultivated areas; pf= Polylepis forest; fi= with out
information. *= endemic species.
Discusión
El
origen de Lachemilla es aún desconocido, Rothmaler
(1937), sugiere un posible origen comun para Alchemilla,
Aphanes y Lachemilla; seria evidente pensar que se
originaron en el hemisferio norte, ya que Alchemilla y Aphanes
son mucho más diversos en el norte. Por otro lado, Kalkman
(1988) propone la hipótesis de que toda la familia Rosaceae se
originó en Gondwana, propiamente en sudamérica y desde
ahí muchos grupos migraron y se diversificaron en el norte, en
este caso Lachemilla se habria originado en el sur; sin
embargo, no hay ningun registro fósil de Rosaceae tan antiguo
que pueda apoyar esta hipótesis.
Actualmente
Lachemilla presenta marcados caracteres morfológicos que
lo diferencian de Alchemilla y Aphanes, y su
distribución sugiere que es un grupo que se desarrolló
en las montañas neotropicales con su centro de diversidad en
los páramos sudamericanos; sin embargo, es importante conocer
su origen, probablemente estudios moleculares podrían ayudar a
aclarar su parentezco con otros taxa y así entender mejor la
evolución y el desarrollo natural de Lachemilla.
Agradecimientos
Agradezco
a la Fundación Alexander von Humboldt por financiar el
proyecto sobre la revisión de Lachemilla y a la
Pontificia Universidad Católica del Ecuador y al Instituto de
Botánica Sistemática de la Universidad Ludwig
Maximilian de Munich por el apoyo académico.
Referencias
Ericksson,
T.; S. Hibbs, A. Yoder, Ch. Delwiche & M. Donoghue. 2003. The
Phylogeny of Rosoideae (Rosaceae) based on sequences of the Internal
Transcribed Spacer (ITS) of nuclear ribosomal DNA and the TRNL/F
region of chloroplast DNA. Ins. J. Plant Sci. 164 (2):
197-211.
Focke,
W. 1888. Rosaceae. En: A. Engler & E. Prantl. (eds.).
Die Natür. Pflanzenfam. III (3): 1-61
Gaviria,
J. 1996. Sinópsis del género Lachemilla (Focke)
Rydberg (Rosaceae) para Venezuela. Plantula.
1 (3): 189-212.
Hutchinson,
J. 1964. The Genera of Flowering Plants I. Pp. 174-216.
Oxford.
Kalkman,
C. 1988. The Phylogeny of the Rosaceae. Bot. J. Linn. Soc. 98:
37-59.
Luteyn,
J. 1999. Páramos: a checklist of plant diversity, geographical
distribution and botanical literature. Memoirs of the New York
Botanical Garten. 84: 1-278.
Perry,
L. 1929. A tentative revision of Alchemilla subgenus
Lachemilla. Contr. Gray Herb. 84: 1-57.
Romoleroux,
K. 1996. Rosaceae. En: Harling, G. & L. Andersson (eds.). Flora
of Ecuador. 56: 1-152.
Rothmaler,
W. 1935. Alchemillae Columbianae. Trab. Mus. Nac. Ci. Nat. Ser.
Bot. 31: 1-52.
Rothmaler,
W. 1937. Systematische Vorarbeiten zu einer Monographie der Gattung
Alchemilla (L.) Scop. emend. VII.
Aufteilung der Gattung und Nomenclatur. Repert. Spec. Nov. Regni
Veg. 42: 164-173.
Rydberg,
A. 1908. Rosaceae. En: Britton & Underw. (eds.). N. Amer. Fl.
22: 377-385.
Schulze-Menz,
G. 1964. Rosaceae. En: A. Engler Syllabus der Pflanzenfamilien,
2 13th ed. By H. Melchior. Pp. 209-218. Berlin.